
Helicops boitata, Moraes-Da-Silva & Amaro & Sales Nunes & Strüssmann & Junior & Jr & Sudré & Recoder & Rodrigues & Curcio, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4651.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:FA2E3659-C2A3-45E9-BBB0-3798EE05808E |
|
persistent identifier |
https://treatment.plazi.org/id/00047545-FFDA-FFEB-AFDA-FCA4EA7FEC76 |
|
treatment provided by |
Plazi |
|
scientific name |
Helicops boitata |
| status |
sp. nov. |
Helicops boitata sp. nov.
( Figs. 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
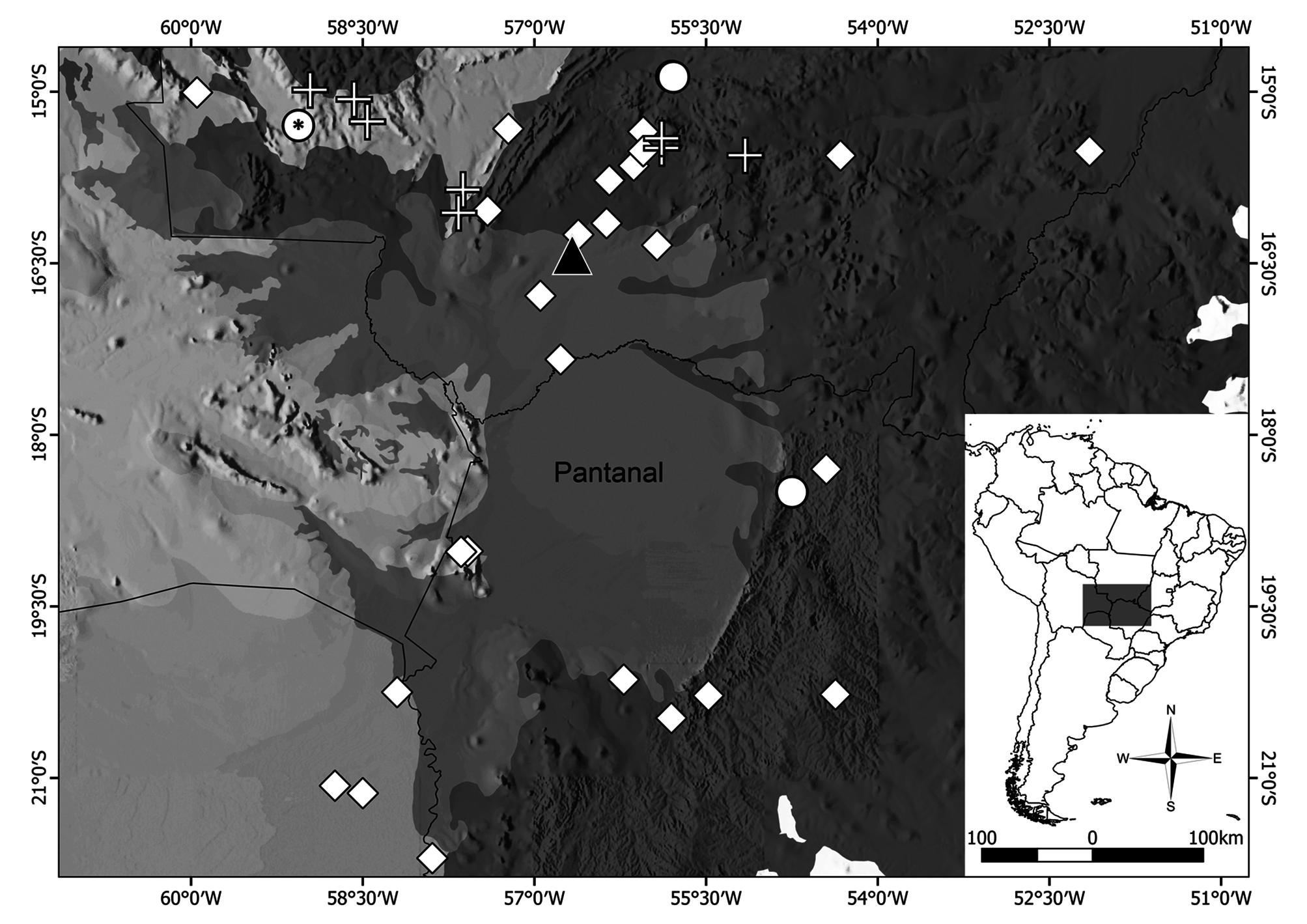
Holotype. An adult male ( UFMT-R 11940 ) collected on May 1 st 2016 by F.F. Curcio; M. Teixeira Jr., R. Recoder and V. Sudré at Transpantaneira Road (16º25’21.18’’S, 56º40’12.64’’W; 124 m above sea level), municipality of Poconé , Mato Grosso state, Brazil ( Fig. 5 View FIGURE 5 ). GoogleMaps
Diagnosis. Helicops boitata differs from all other congeners for presenting the unique combination of the following characters: (1) dorsal scale rows 25/25/21, with moderate keels throughout the whole body length, slightly stronger on posterior one-third of trunk; (2) low ventral counts (113, male); (3) moderate subcaudal counts (68, male; tail tip missing); (4) subcaudal keels absent, moderate keels present on tail dorsals; (5) supralabials 10/ 10, 4 th– 6 th contacting orbit; (6) infralabials 12/13; (7) intergenials absent; (8) nasal entire; (9) two preoculars; (10) two postoculars; (11) temporals 1+3; (12) maxillary teeth 20+2; (13) hemipenis strongly bilobed, noncalyculate, hemicapitate with a shallow capitular groove, lobes ornamented with circular spinulate transverse folds, body elongate, nearly twice as long as lobes, ornamented with enlarged lateral spines and small spicules in the central area of asulcate face; (14) dorsum greenish copper brown, with one vertebral and two lateral chain-like rows composed of dark spots, ventrally flanked by a continuous light orange paraventral stripe, and (15) venter light brown, with two irregular stripes composed of light orange rounded spots along the lateral margins of ventrals that converge towards midline of the ventral surface of tail, ending at level of 38 th subcaudal.
Comparisons. The most relevant features that distinguish Helicops boitata from other congeners refer to the presence of entire nasal plate and the general color pattern.An entire nasal plate occurs only in H. nentur , H. petersi , and H. yacu (but see Remarks for comments on H. petersi ). The presence of entire cloacal plate in the holotype of H. boitata (see Description of the holotype) appears as exclusive to the new species in the genus, although we concede that our restriction to one single specimen renders the uniqueness of this feature dependent on further sampling. For this reason, we decided to exclude the condition of the anal plate from our diagnosis until new specimens come to clarify this issue.
Regarding color pattern, there are no other Helicops species with a ventral pattern similar to the one of Helicops boitata , with a light brown venter ornamented by vivid orange rounded spots forming irregular ventral stripes ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 ). The dorsal pattern in life is also unique; no other congener exhibits the chain-like vertebral and lateral spot rows combined with the continuous paraventral light orange stripe, eliminating any possibility of misidentification. Dorsal patterns documented in Helicops taxa are mostly composed of alternated spots/blotches ( H. danieli , H. gomesi , H. hagmanni , H. leopardinus , H. pastazae , H. petersi , H. polylepis , H. scalaris , and H. yacu ), with a lower number of representatives with a uniform dorsum ( H. nentur and H. tapajonicus ), dorsal bands ( H. angulatus , H. apiaka ), or stripes ( H. carinicaudus , H. infrataeniatus , H. modestus , and H. trivittatus ) (see Costa et al. 2016).
The elongate and slender hemipenial lobes promptly distinguish Helicops boitata from H. angulatus , H. apiaka , H. carinicaudus , H. danieli , H. gomesi , H. infrataeniatus , H. leopardinus , H. modestus , H. polylepis , and H. tapajonicus (vs. short or globose hemipenial lobes); the presence of enlarged lateral spines on the hemipenial body distinguishes the new species from H. hagmanni , H. infrataeniatus , H. leopardinus , H. modestus , H. tapajonicus , and H. trivittatus (vs. enlarged lateral spines absent); the elongate and slender hemipenial body distinguishes the new species from H. angulatus , H. apiaka , H. carinicaudus , H. danieli , H. gomesi , H. infrataeniatus , H. leopardinus , H. modestus , H. polylepis , and H. tapajonicus (vs. globose or cylindrical hemipenial body); an hemicapitate hemipenis with a shallow capitular groove distinguishes the new species from H. angulatus , H. apiaka , H. carinicaudus , H. danieli , H. gomesi , H. pastazae , H. polylepis , and H. tapajonicus (vs. hemipenis bicapitate with a deep capitular groove); the absence of reduced hook-shaped calcified spines dispersed through the central area of the asulcate face distinguishes the new species from H. hagmanni , H. infrataeniatus , H. leopardinus , H. modestus , H. pastazae , H. tapajonicus , and H. trivittatus , in which such structures are present.
The 25/25/21 dorsal rows of Helicops boitata exceed at least the dorsal formulae of H. nentur (17/17/15), H. carinicaudus (19/17–19/17–19), H. infrataeniatus (17–19/19/17–19), H. angulatus , H. modestus , H. tapajonicus (19/19/17), H. gomesi (19/19/17–19), H. danieli (19/19/19), H. leopardinus (19–20/19/17–19), H. apiaka (21– 23/19–21/19), H. petersi (21/21–23/16), and H. pastazae (23/21–23/19). The only comparable taxa that may possibly exhibit 25 dorsal rows at some region of the trunk are H. hagmanni (27/23–27/23), H. polylepis (23–25/22– 25/19–21), H. scalaris (21-25/19-21/16-19), H. trivittatus (21–25/21–23/21–19), and H. yacu (25–29/25–28/18–20) ( Table 4 View TABLE 4 ).
According to our definition, Helicops boitata exhibits moderate dorsal keels¸ similar to the patterns present in H. danieli , H. infrataeniatus , H. leopardinus , and H. trivittatus . Diverge from such a condition the taxa H. angulatus , H. apiaka , H. gomesi , H. hagmanni , H. pastazae , H. petersi , H. polylepis , and H. scalaris (strong dorsal keels), as well as H. modestus , H. nentur , and H. tapajonicus (weak dorsal keels). Prominence of keels vary between weak and moderate in H. carinicaudus ( Table 4 View TABLE 4 ). Helicops boitata lacks any traces of subcaudal keels, rendering it promptly distinguishable from H. angulatus , H. apiaka , and H. gomesi (prominent subcaudal keels) and H. scalaris (moderate subcaudal keels) (see Table 4 View TABLE 4 ). The literature also reports such a feature in H. infrataeniatus (table 1 in Kawashita-Ribeiro et al. 2013:81), but this information is inconsistent and requires revision, as our data on this species reveal no evident subcaudal keels, as also reported by Costa et al. (2016:161, table 3).
Helicops boitata has the highest number of supralabials in the genus (10/10). Our data revealed no more than nine supralabials in all species from which we had primary data, whereas the counts provided in the original descriptions of H. scalaris (nine supralabials) and H. yacu (nine) are also below this limit ( Table 4 View TABLE 4 ). Literature reports 10 supralabials to H. leopardinus and H. pastazae ( Kawashita-Ribeiro et al. 2013; Costa et al. 2016), but we argue that such information deserves checking (see Remarks). The holotype of Helicops boitata has 12/13 infralabials (lateral asymmetry), also reflecting high counts for this character within the genus, only comparable to those from H. apiaka , H. hagmanni , H. petersi , H. yacu (up to 12), H. pastazae , H. polylepis (up to 13), H. trivittatus , and H. scalaris (up to 14). The upper bound of infralabials of the remaining congeners is always below these limits, with counts of up to 11 scales in H. angulatus , H. danieli , H. gomesi , H. infrataeniatus and H. modestus , and up to 10 scales in H. carinicaudus , H. leopardinus , H, nentur , H. pastazae , and H. tapajonicus ( Table 4 View TABLE 4 ).
Finally, the last relevant character that may help in the distinction of Helicops boitata refers to the two preoculars, similar to H. angulatus , H. carinicaudus , H. hagmanni , H. leopardinus (vs. one or two), and H. trivittatus (vs. two). All other congeners have only one preocular scale ( Table 4 View TABLE 4 ).
Description of the holotype ( Figs. 2–3 View FIGURE 2 View FIGURE 3 ). Body moderately stout (midbody diameter/ SVL = 0.03); total length 642 mm ( SVL 437 mm, tail length 205 mm); head length 22.64 mm; head width 12.64 mm; snout length 6.28 mm; snout width 5.60 mm; right eye diameter 2.53 mm; eye-nostril distance 4.10 mm; neck width 8.40; diameter at midbody 14.53 mm; tail length almost one-third of total length (tail length/total length = 0.32); head elongated, arrow-shaped, almost twice as long as wide (head width/head length = 0.56), distinct from neck (neck width/head width = 0.67); snout slightly longer than wide (snout width/snout length = 0.89), acuminate in dorsal view, its length corresponding to slightly more than one-fourth of head length (snout length/head length = 0.28); eyes relatively small, slightly displaced dorsally; eye diameter much shorter than head length (eye diameter/head length = 0.11) and slightly longer than one-half the distance from its anterior margin to nostril center (eye diameter/eyenostril distance = 0.62); pupil rounded; rostral feebly pointed, not projecting over lower jaw, widely visible from above; internasal single, roughly triangular, slightly longer than wide, lateral margins feebly concave, posterior margin convex; internasal contacting tip of rostral, nasals and prefrontals; prefrontals paired, roughly pentagonal, posterior border slightly convex; prefrontals wider than long, largest length nearly one-third of largest width, and distinctly larger than length of their mutual suture; frontal pentagonal, twice as long as wide, its lateral borders slightly convex; frontal in contact with prefrontals, supraoculars, and parietals; supraoculars large, with five borders, irregular in shape, nearly twice as long as wide, in contact with upper preoculars, prefrontals, frontal, parietal, upper postocular, and dorsal margin of eye; parietals paired, longer than wide, largest width nearly two-thirds of largest length, and slightly longer than their mutual suture; lateral border of right parietal notched at level of dorsal vertex of upper primary temporal; medial border of left parietal notched at midpoint and rear extremity of parietal suture; parietals contacting frontal, supraoculars, upper postocular, primary, and upper secondary temporals and anteriormost dorsals; nasal entire, roughly semielliptical, in contact with rostral, internasal, prefrontal, loreal, and 1–2 nd supralabials; nostril positioned on central area of nasal, totally displaced dorsally, entirely visible from above; nostrils equipped with flap-like nasal valves, anteromedially directed and connected to posterior margin of external naris; loreal pentagonal; nearly as high as long, in contact with nasal, prefrontal, preoculars, and 2–3 rd supralabials; oculars 2+2; upper preocular pentagonal, slightly longer than high, in contact with loreal, prefrontal, supraocular, lower preocular, and anterior margin of eye; lower preocular trapezoidal, longer than high; its largest height slightly larger than two-thirds its largest length, in contact with loreal, upper preocular, 3–4 th supralabials, and anterior margin of eye; upper postocular roughly trapezoidal, higher than long, its largest length nearly two-thirds its largest height, in contact with parietal, primary temporal, lower postocular, and posterior margin of eye; lower postocular roughly pentagonal, longer than high, its largest height nearly two-thirds its largest length, in contact with upper postocular, primary temporal, 6–7 th supralabials, and posterior margin of eye; temporals asymmetrical, 2+2+3 (right side), 1+2+3 (left side), most roughly pentagonal, decreasing in size from primary temporals backwards; supralabials 10/10, 4–6 th in contact with the orbit, 7–9 th largest; 7–10 th supralabials separated from parietals by temporals; mental triangular, slightly longer than wide; two pairs of chinshields, posterior pair slightly longer than anterior pair; infralabials 12/13 (lateral asymmetry), 1–6 th (right side) and 1–7 th (left side, only anteromedial tip of 7 th scale) contacting anterior chinshields, 6–8 th (right side) and 7–9 th (left side) contacting posterior chinshields; 8 th (right side) and 9 th (left side) infralabials largest, roughly pentagonal, only the medial tip contacting second chinshield on each side of head; 1 st pair of infralabials in contact posteriorly to mental; dorsals 25/25/21, mostly keeled, without apical pits; dorsal keels weak on anterior one-third of trunk, increasing backwards in prominence and becoming moderate from midbody to tail; keels sensibly weaker on rows closer to venter, mostly absent on paraventral dorsal scales of anterior two-thirds of trunk; cloacal plate entire; ventrals 113, first twelve anomalously divided, except for 8 th one; subcaudals 68/68, tail tip broken slightly anteriorly from terminal scale; maxillary teeth 20+2, 9 th and 16 th missing, diastema present; prediastemal teeth slightly increasing in size backwards; postdiastemal teeth ungrooved, slightly larger than prediastemal ones.
Coloration in preservative ( Figs. 2–3 View FIGURE 2 View FIGURE 3 ). Head dorsum mostly uniform dark greenish brown, with two transversal slightly darker spots on anterior portion of supraoculars, and an irregular and weakly visible dark ocular stripe; most upper area of supralabials slightly lighter than head dorsum, with barely visible darker marks on lower and/or lower anterior surfaces of scales, as well as minute light pale dots on ventral margin of most scales; infralabials and ventral surface of head salt-and-pepper patterned, slightly darker at level of infralabials; infralabials ornamented with light pale dots present at least from 5–10 th scales; pupil black; iris mostly light greyish brown, reticulated in a marble texture; iris with two brown triangular spots anterior and posterior to pupil, aligned with irregular and barely visible ocular stripe; body dorsum greenish copper brown; dorsum with three chain-like longitudinal rows of dark spots; vertebral spot row starting at nape level, almost indiscernible from background; vertebral spots irregular in shape barely connected to each other or separated from each other by no more than one-half scale; vertebral spots becoming gradually narrow backwards, and assuming a longitudinal slit shape on tail; lateral spot row starting at neck level, more distinct from background from level of 12 th ventral backwards; lateral spots of trunk 54 on right side, 56 on left side, roughly rectangular, slightly longer than wide, covering 5–8 th rows of dorsals (not completely) and two-and-a-half scales in length at midbody; lateral spots of dorsum feebly connected to each other, abruptly decreasing in size backwards at level of 100 th ventral; lateral spots on tail irregular in shape, gradually decreasing in size backwards until tail tip; paraventral dorsal rows pigmented by greyish brown (from 1 st to lower margin of 3 rd row) and light orange (most of 3 rd and lower half of 4 th rows), configuring two continuous and irregular paraventral stripes throughout the whole trunk and almost whole tail length; central area of venter mostly light greyish brown, flanked by subsequential roughly rounded light orange spots covering most of lateral margins of ventral scutes and some of the scales of first dorsal row; some light orange spots also present on central area of some ventrals, especially in posterior two-thirds of trunk; on ventral surface of tail, orange spots abruptly converge to midline, proceeding backwards until level of 35 th subcaudal; background of ventral surface of tail light greyish brown, following tonality of central area of venter.
Coloration in life ( Fig. 4 View FIGURE 4 ). Quite similar to the condition described in preserved state, although head and body markings are more clearly visible in life due to the contrast between the lighter background tonality and dark spots. Head spots visible on internasal, oculars, prefrontals and frontal (the latter forming a transversal irregular band slightly anterior from eye level) are virtually indiscernible in preservative, and the barely visible irregular ocular stripe and dark spots of supralabials are more evident in the alive holotype. In life, the infralabials appear also marked by the same dark spots present on supralabials, although one cannot distinguish the limits of such structures in the preserved specimen. Photographs of the lateral surface of head of the alive holotype also suggest that the lateral quadrangular spots of dorsum represent the fusion of the dark markings of labials that converge in the corner of the mouth and assume a more evident conformation on the neck and backwards. Finally, the light orange markings present on paraventrals, ventrals, and subcaudals are vivid reddish orange in life.
Hemipenial morphology ( Fig. 6 View FIGURE 6 ). Description based on the fully everted and expanded right organ of the holotype. Inverted organ 11 subcaudals in length, distal end of retractor magnus divided. Hemipenis strongly bilobed, noncalyculate, hemicapitate, with a shallow capitular groove; body conical, elongate, almost two-thirds the length of organ; lobes elongate, well developed, roughly cylindrical, strongly divergent, nearly one-third the length of organ, without apical disks; base nude, nearly one-fourth the length of body, small basal pocket present, dextral from perspective of asulcate face; body ornamented by a row with five enlarged hook-shaped spines on each flank of asulcate face, increasing in size from base to lobes; sulcate face ornamented by smaller hook-shaped spines dispersed towards body flanks; body spines interspersed by small spicules especially concentrated in central area of asulcate face; lobes encircled by 8–9 prominent spinulate flounces; sulcus spermaticus shallow, strongly bifurcate, centrifugally oriented, dividing at proximal one-third of body, running throughout the median surface at the lobe base and progressively assuming a lateral orientation to end laterally at lobe tip.
Natural history and habitat ( Fig. 7 View FIGURE 7 ). We found the holotype at 14:23hs, almost entirely submerged in a flooded area at the margins of the “Transpantaneira”, an unpaved road crossing the northern portion of Pantanal wetlands. This finding suggests that Helicops boitata has diurnal habits, contrasting with other congeners considered nocturnal ( H. angulatus and H. polylepis ) ( Marques et al. 2005). In the Pantanal, at least H. leopardinus may be active during the day and night ( Marques et al. 2005). Detecting the specimen became easy because it was exposed to the sun and the water in the collection spot was clear, shallow (up to 30 cm deep), and free from floating aquatic plants that would otherwise obstruct the vision. The collection site corresponds to a tributary of the Bento Gomes River that spreads in the area during the rainy season (from November to February), receding back to the riverbed in the dry season (from June to August) ( Cunha & Junk 2009:305). Thus, the collection date reflects the receding period, when flooded areas start to dry out. We visited again the collection site in July of 2018–dry season–and the place still retained water in isolated and very turbid ponds.
The specimen did not bite when manipulated, contrasting with the usually aggressive behavior of most congeners that promptly bite when grabbed ( Marques et al. 2005). Spectacled caimans [ Caiman yacare ( Daudin, 1802)] are abundant in the area, as well as many bird species such as herons ( Ardea alba , Egretta thula , and Tigrisoma lineatum ; Ardeidae ), storks ( Mycteria americana , Jabiru mycteria ; Ciconidae ) and hawks ( Busarellus nigricollis ; Accipitridae ) that may prey on snakes ( Sick 1997). In the same day we collected the holotype, we also found one specimen of Helicops leopardinus on the opposite margin of the road, a few steps from the site where we collected the holotype of H. boitata .
Etymology. The specific name is a noun in apposition deriving from the Brazilian Portuguese “boitatá”, as a reference to the folk legend of a giant fire snake that inhabits the Brazilian rivers and protects the forests against man-caused fire. The word has its origins in the native language Tupi (‘Mboi = snake; tatá = fire). It serves also as an allusion to the vivid orange tonality of the ventral pattern, resembling the color of flames.
Remarks. The literature on Helicops taxonomy brings contradictory pieces of information that only the objective examination of specimens can elucidate. For instance, the entire condition of the nasal plate alluded in the original description of H. petersi confronts the data reported by Costa et al. (2016), as well as the data obtained from a collaborator who directly examined the holotype (see Acknowledgements). This is why we decided to restrict our comparisons to the specimens from our sample, conceding that data retrieved from vouchers examined in recent taxonomic studies ( Kawashita-Ribeiro et al. 2013; Costa et al. 2016) should also be considered.
Our comparative material comprises 16 of the 17 currently recognized species of Helicops . In that sense, our study represents the most comprehensive taxonomic approach of the genus Helicops until present. Thus, we were able to confirm all the morphological traits at least for these 16 taxa through direct observation of specimens, while our information regarding H. yacu remained restricted to literature data. In addition, our phylogenetic approach is also the most complete in terms of diversity, including nine Helicops taxa besides the new species [vs. six Helicops terminals in Costa et al. (2016)].
The high number of supralabials of Helicops boitata (10/10) represents an informative diagnostic feature. The only taxa with the same number of supralabials would be H. leopardinus and H. pastazae (sensu Kawashita-Ribeiro et al. 2013 and Costa et al. 2016). However, our data based on vouchers of both species do not reflect the varia- tions reported in the literature. For instance, while our vouchers of H. pastazae ( MZUSP 6111, 14108) agree with the original description ( Shreve 1934:129) in having both eight supralabials, literature data report up to 10 scales in the same species ( Rossman 1976:15; Kawashita-Ribeiro et al. 2013:82; and Costa et al. 2016:161). Nevertheless, Kawashita-Ribeiro et al. (2013) and Costa et al. (2016) had not personally examined vouchers, only replicating the data of Rossman (1976). Regarding H. leopardinus , our vouchers (45 specimens) have all nine supralabials, whereas the larger sample of Kawashita-Ribeiro et al. (2013) —259 specimens—includes at least some individuals with 10 scales. In both cases, we assume that supralabial counts are subjected to variation, although further revisions must throw light on the precise levels of polymorphisms, as well as on their taxonomic significance.
The unique coloration features of Helicops boitata appear to represent an autapomorphic condition, not comparable with patterns of any other congener. Finally, the entire cloacal plate of the holotype diverges from all Helicops taxa; nonetheless, although it may also represent a diagnostic feature, we refrained to include it in the formal diagnosis until the discovery of additional specimens allow more conclusive decisions regarding the generality of this character state.





| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Helicops boitata
| Moraes-Da-Silva, Antonio, Amaro, Renata Cecília, Sales Nunes, Pedro M., Strüssmann, Christine, Junior, Mauro Teixeira, Jr, Albedi Andrade, Sudré, Vinícius, Recoder, Renato, Rodrigues, Miguel Trefaut & Curcio, Felipe Franco 2019 |
Helicops boitata
| Moraes-Da-Silva & Amaro & Sales Nunes & Strüssmann & Junior & Jr & Sudré & Recoder & Rodrigues & Curcio 2019 |
Helicops boitata
| Moraes-Da-Silva & Amaro & Sales Nunes & Strüssmann & Junior & Jr & Sudré & Recoder & Rodrigues & Curcio 2019 |
Helicops boitata
| Moraes-Da-Silva & Amaro & Sales Nunes & Strüssmann & Junior & Jr & Sudré & Recoder & Rodrigues & Curcio 2019 |
nentur
| Costa, Santana, Leal, Koroiva & Garcia 2016 |
H. apiaka
| Kawashita-Ribeiro, Avila & Morais 2013 |
H. tapajonicus
| Da Frota 2005 |
H. petersi
| Rossman 1976 |
H. yacu
| Rossman & Dixon 1975 |
H. yacu
| Rossman & Dixon 1975 |
H. danieli
| Amaral 1938 |
H. pastazae
| Shreve 1934 |
H. pastazae
| Shreve 1934 |
H. pastazae
| Shreve 1934 |
H. gomesi
| Amaral 1921 |
H. hagmanni
| Roux 1910 |
H. hagmanni
| Roux 1910 |
H. trivittatus
| Cope 1878 |
H. trivittatus
| Cope 1878 |
H. infrataeniatus
| Jan 1865 |
H. polylepis
| Guenther 1861 |
H. modestus
| Guenther 1861 |