
Lycenchelys squamosa Toyoshima, 1983
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4762.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:BEBD8F0D-1347-4A44-86D4-2915433D2E7B |
|
DOI |
https://doi.org/10.5281/zenodo.3809763 |
|
persistent identifier |
https://treatment.plazi.org/id/006C5E1A-FFBD-FFAD-3EC6-B2C8FCC3AE4C |
|
treatment provided by |
Plazi |
|
scientific name |
Lycenchelys squamosa Toyoshima, 1983 |
| status |
|
Lycenchelys squamosa Toyoshima, 1983 View in CoL
(Japanese name: Uroko-hebigenge)
( Figs. 38–43 View FIGURE 38 View FIGURE 39 View FIGURE 40 View FIGURE 41 View FIGURE 42 View FIGURE 43 ; Table 10)
Lycenchelys squamosus Toyoshima, 1983: 145 View in CoL , figs. 20–22, table 10, pl. 93 (original description, type locality: off Miyagi Prefecture, Pacific coast of Honshu Island, Japan); Toyoshima, 1984: 293, pl. 274-E (brief description); Toyoshma, 1985: 156, figs. 6–7, 12–13, 31, table 1 (description); Hatooka, 1993: 901, unnumbered fig. (key to species); Amaoka et al., 1995: 241, pl. 405 (brief description); Koyanagi, 1997: 538, fig. 5 (brief description); Imamura, 1998: 32, fig. 13 (brief description); Zama, 2001: 86, 133 (species list).
Lycenchelys squamosa: Anderson, 1994: 113 View in CoL , 118 (species list); Shinohara et al., 1996: 182 (species list); Imamura, 1997: 60 (species list); Shinohara & Matsuura, 1998: table 1 (comparison with Lycenchelys aurantiaca ); Hatooka, 2000: 1033, unnumbered fig. (key to species); Hatooka, 2002: 1033, unnumbered fig. (key to species); Anderson & Fedorov, 2004: 20 (species list); Shiogaki et al., 2004: 71 (species list); Shinohara & Anderson, 2007: 64 (key to species); Kitagawa et al., 2008: 96, unnumbered fig. (brief description); Shinohara et al., 2009: 724 (species list); Amaoka et al., 2011: 318, unnumbered fig. (brief description); Balushkin et al., 2011: 983 (catalog of specimens); Hatooka, 2013: 1227, unnumbered fig. (key to species); Nakabo & Hirashima, 2015: 217 (species list and etymology of scientific name).
Materials examined
Holotype: HUMZ 78464 , male, 246.1 mm SL, off Miyagi Prefecture, Tohoku District, northwestern Pacific (37°49.7’N, 142°23.7’E to 37°55.4’N, 142°24.5’E), 985–1005 m depth, 8 Oct. 1978. GoogleMaps
Paratypes (2 specimens, 193.0– 222.5 mm SL, both from Tohoku District, northwestern Pacific): HUMZ 72563, 1 female, 193.0 mm SL, off Fukushima Prefecture (37°09.4’N, 141°56E), 900–920 m depth, 19 Jan. 1978; HUMZ 78390, 1 female, 222.4 mm SL, off Aomori Prefecture (40°47.6’N, 142°16.7’E to 40°41.5’N, 142°19.2’E), 920–948 m depth, 11 Sep. 1978.
Other specimens (6 specimens, 203.0– 253.1 mm SL): HUMZ 157660, 157669, 181894–95, 192737, 2 males and 3 females, 203.0– 253.1 mm SL, Tohoku District, northwestern Pacific; HUMZ 177036, 1 female, 241.8 mm SL, eastern Hokkaido Island, northwestern Pacific.
Diagnosis. Vertebrae 20 + 71–75 = 91–95; head length 16.3–19.9% SL; interorbital pore 1; occipital pores absent; postorbital pores 4; suborbital pores 5 + 1 (rarely 5 + 2); preoperculomandibular pores 8; vomerine teeth 7–17; palatine teeth 5–25, arranged in 1 or 1–2 rows; opercular flap well-developed; pelvic-fin base positioned below lower edge of gill opening; lateral line incomplete and positioned ventrally; scales present on pectoral fin and its base; body uniformly dark brown when fresh.
Description. Counts and proportional measurements in Table 10.
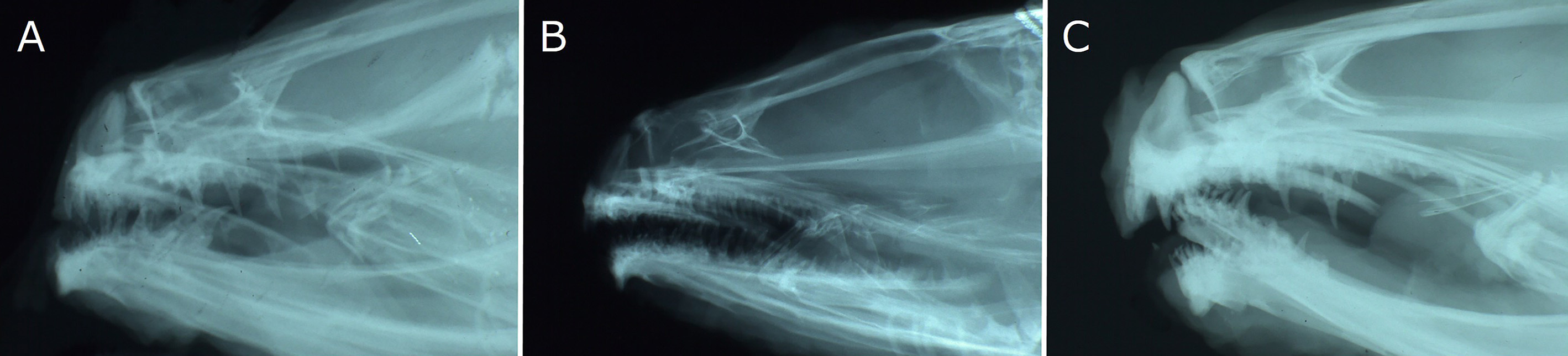
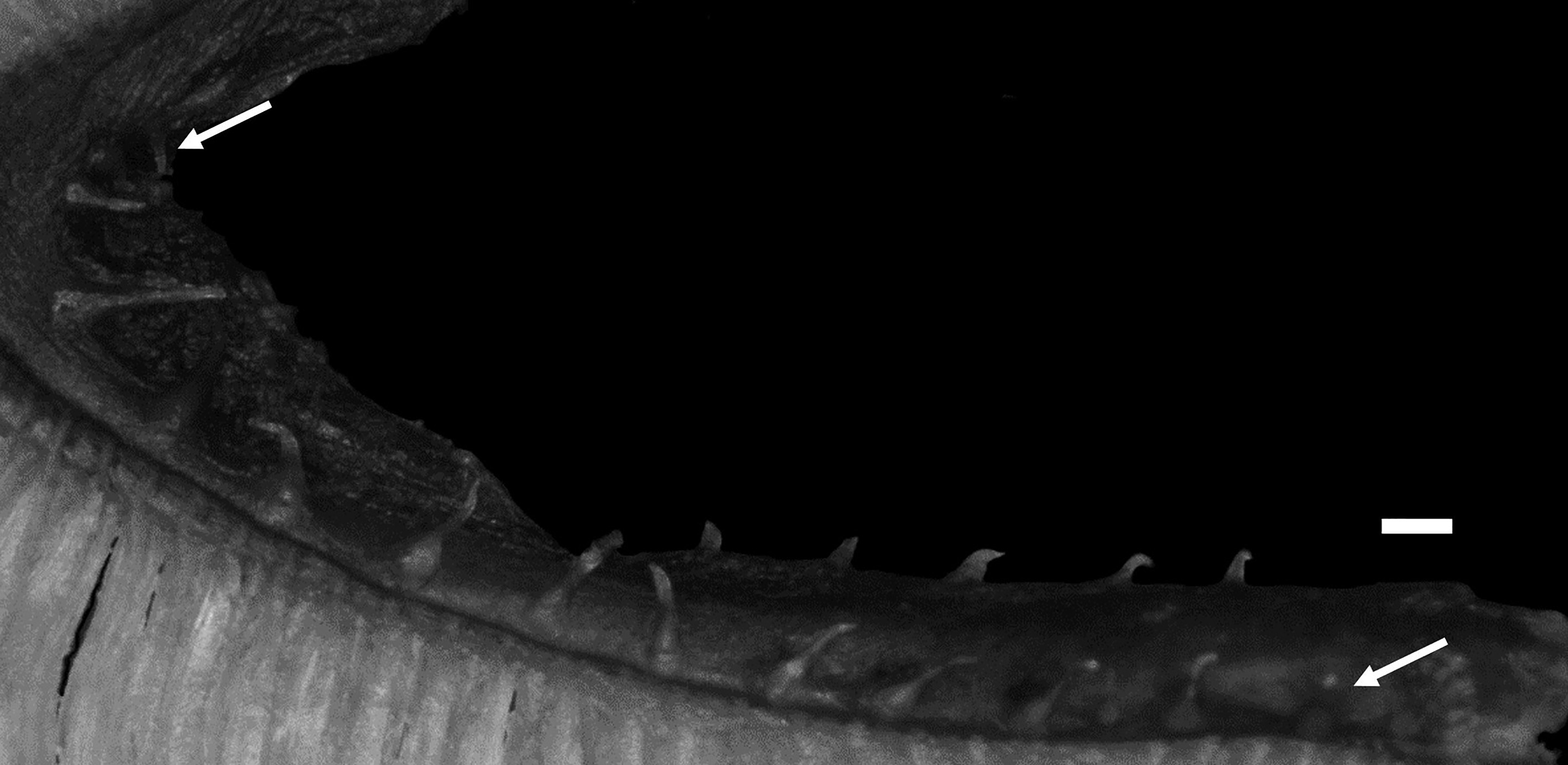
Body elongate, cross section oval anteriorly, compressed laterally near tail; its width at anal-fin origin 5.4–6.5 (5.7)% SL. Head moderately long, ovoid; dorsal profile of head gently sloping to dorsal-fin origin. Snout short, 125.8–173.2 (159.3)% of eye diameter. Eye ovoid, relatively large. Interorbital space narrow, width 15.6–35.6 (27.4)% of eye diameter. Nostril tube short, not reaching upper lip when depressed. Mouth subterminal; lower jaw extending slightly beyond upper jaw (holotype) ( Fig. 39A View FIGURE 39 ), or upper jaw extending slightly (paratypes and non-type specimens, all females) or significantly (non-type males) beyond lower jaw ( Fig. 39B, C View FIGURE 39 ). Posterior end of upper jaw reaching to or slightly beyond vertical through suborbital pore behind eye in males, reaching below posterior margin of eye in females. Labial lobe of lower jaw weak. Teeth on jaws, vomer and palatine small and conical; upper jaw with 2–3 rows anteriorly and single row posteriorly; lower jaw with 2–4 irregular rows anteriorly and single row posteriorly; vomerine teeth irregularly arranged; all palatine teeth in single row, or in 2 rows anteriorly and single row posteriorly (single row). Lower edge of gill opening slightly above lower end of pectoral-fin base. Opercular flap well developed. Gill rakers short and thin ( Fig. 40 View FIGURE 40 ). Pseudobranch filaments short. Lateral line deciduous, positioned ventrally and incomplete; its origin posterior to last postorbital pore and terminating above middle of anal fin. Scales small and cycloid, present on nape, body, pectoral axilla, about basal half of pectoral fin, pectoral-fin base, tail and vertical fins except at margin. Head without scales.
Dorsal-fin origin near vertical through posterior edge of opercular flap; 1st dorsal-fin pterygiophore between neural spines of 2nd and 3rd vertebrae. Anal-fin origin below 18th to 20th (19th) dorsal-fin ray; 1st anal-fin pterygiophore posterior to parapophysis of ultimate abdominal vertebra or hemal spine of 1st caudal vertebra (posterior to parapophysis of ultimate abdominal vertebra). Last dorsal-fin pterygiophore between neural spines of 2nd to 5th (between 4th and 5th) preural vertebrae. Last anal-fin pterygiophore between hemal spines of 2nd to 5th (between 3rd and 4th) preural vertebrae. Caudal fin with 1–2 (2) epural, 4 upper hypural and 3–4 (4) lower hypural rays. Pectoral fin relatively long, reaching to about mid-portion of abdomen; its posterior margin rounded. Upper end of pectoral-fin base on about lateral midline of body. Pelvic fin short; its posterior margin slightly beyond lower edge of gill opening.
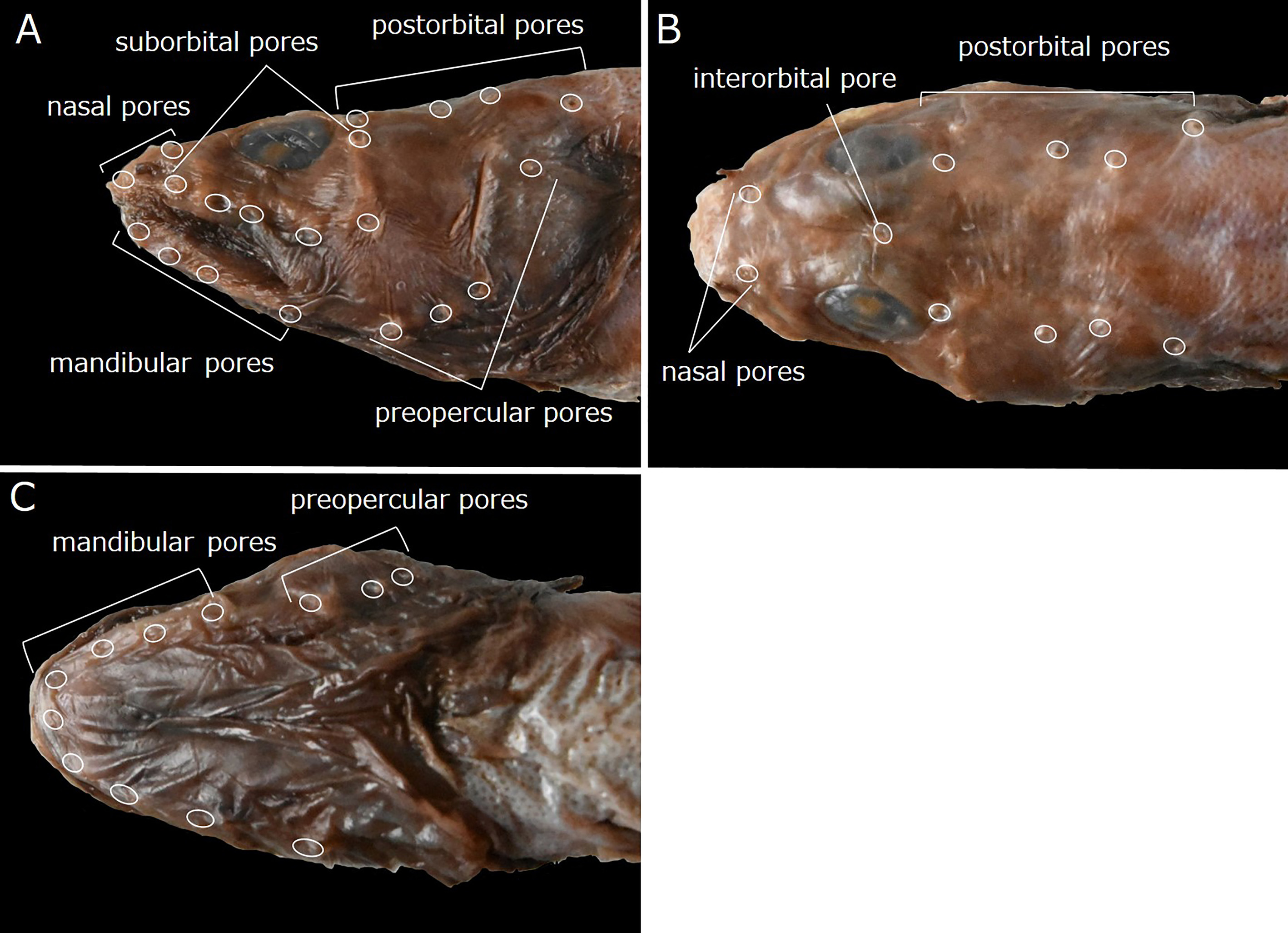
Head pores well developed and distinct. Nasal pores 2; anterior pore in front of nostril tube, posterior pore above and slightly anterior to vertical through 1st suborbital pore ( Fig. 42A, B View FIGURE 42 ). Postorbital pores 4; distance between 1st and 2nd pores longest of those between adjacent pores; 3rd pore slightly higher than others ( Fig. 42A, B View FIGURE 42 ). Suborbital pores usually 6 (including holotype), rarely 7; 5 pores below eye and last pore behind midline of eye, except for another pore just above 5th pore on left side in HUMZ 181895 and right side in HUMZ 157669; 5th pore behind vertical through 1st postorbital pore ( Fig. 42A View FIGURE 42 ). Preoperculomandibular pores 8; 4 on lower jaw and 4 on preopercle; last preoperculomandibular pore posterior to lower margin of eye ( Fig. 42A, C View FIGURE 42 ). One interorbital pore on dorsal midline above middle of eyes ( Fig. 42B View FIGURE 42 ). Occipital pores absent ( Fig. 42B View FIGURE 42 ).
Color in alcohol. Holotype ( Fig. 41 View FIGURE 41 ) with light purplish brown head, body and vertical fins, color fading somewhat on body, right side of tail and vertical fins. Pectoral fin slightly lighter than body color. Head, body, vertical and pectoral fins dark brown in paratypes. Head and pectoral fin dark brown, body and vertical fins grayish brown, margins of dorsal and anal fins blackish in other specimens.
Color when fresh (based on color photograph of HUMZ 192737; Fig. 38 View FIGURE 38 ). Head and pectoral fin blackish. Body, vertical fins and pectoral-fin base uniformly dark brown. Margins of dorsal and anal fins black.
Distribution. The Okhotsk Sea, off northwestern Pacific coast of eastern Hokkaido Island and in the northwestern Pacific from Aomori to Ibaraki prefectures, at depths of 310–1340 m ( Toyoshima, 1983, 1984, 1985; Hatooka, 1993, 2000, 2002, 2013; Anderson, 1994; Amaoka et al., 1995, 2011; Shinohara et al., 1996; Imamura, 1997, 1998; Koyanagi, 1997; Shinohara & Matsuura, 1998; Anderson & Fedorov, 2004; Shiogaki et al., 2004; Shinohara & Anderson, 2007; Kitagawa et al., 2008; Shinohara et al., 2009; Balushkin et al., 2011; this study).
Size. The largest specimen examined during this study measured 253.1 mm SL (261.3 mm TL), about equal to the previously recorded maximum length (252 mm SL, 260 mm TL) ( Toyoshima, 1983, 1984, 1985; Hatooka, 1993, 2000, 2002, 2013; Anderson, 1994; Amaoka et al., 1995, 2011; Koyanagi, 1997; Shinohara & Matsuura, 1998; Shinohara & Anderson, 2007).
Remarks. Lycenchelys squamosa resembles L. aurantiaca in having less than 100 total vertebrae, 1 interorbital pore, no occipital pores, 4 postorbital pores and no distinct spots or blotches on the body (vs. lacking this combination of characters in other species of Lycenchelys ) (e.g., Toyoshima, 1983, 1985; Shinohara & Matsuura, 1998; Hatooka, 2000, 2002, 2013; Shinohara & Anderson, 2007; this study). See Remarks under L. aurantiaca for a detailed comparison of L. squamosa with L. aurantiaca .
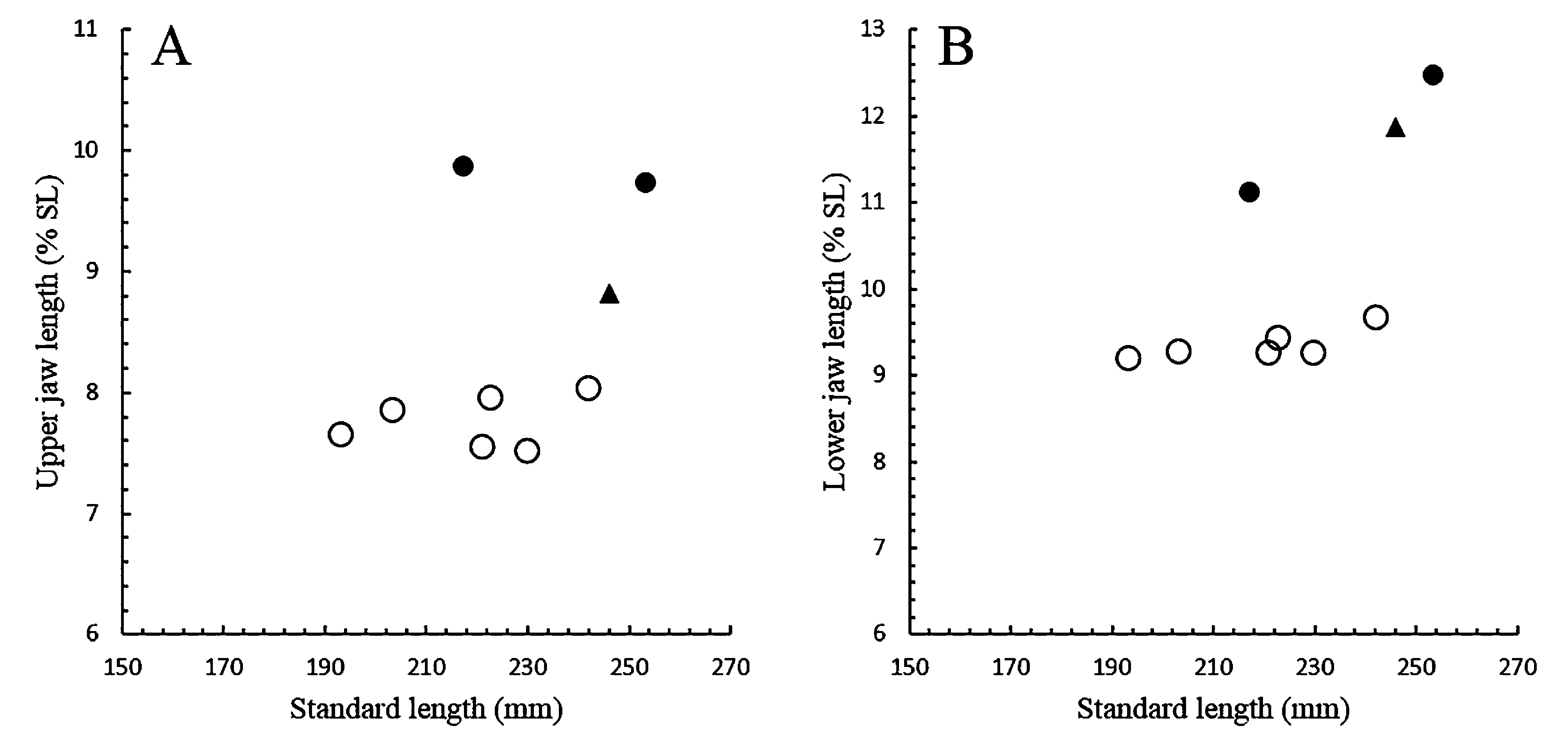
In Lycenchelys squamosa , the upper jaw extends forward slightly beyond the lower jaw in females (including paratypes) ( Fig. 39B View FIGURE 39 ). In males however, the lower jaw extends slightly (holotype, 246.1 mm SL) ( Fig. 39A View FIGURE 39 ) or considerably beyond the lower jaw (2 non-types, 217.0 and 253.1 mm SL) ( Fig. 39C View FIGURE 39 ). According to Toyoshima (1983, 1985), the lower jaw of the holotype (male) protrudes slightly beyond the upper jaw, and the lower jaw is nearly equal to or slightly included in the upper jaw of paratypes (females). He did not mention the variation in the jaw protrusion of males because he was able to examine only 1 male specimen (= holotype). Subsequent studies have not described the variation in males of the species even though the extension of the lower jaw in males of the species as presented by Toyoshima (1985) has been repeated in some studies (e.g., Amaoka et al., 1995, 2011). This study found that the variation in males lies in the length of the upper jaw, not of the lower jaw, although the ratio of the lower jaw length seems to increase with growth ( Fig. 43B View FIGURE 43 ). In the 3 males examined, the 246.1 mm SL specimen (holotype) with the lower jaw protrusion has a shorter upper jaw, while the largest (253.1 mm SL) and smallest (217.0 mm SL) specimens with the upper jaw protrusion have a longer upper jaw ( Fig. 43A View FIGURE 43 ). Therefore, the length of the upper jaw may be due to intraspecific variation not associated with change in growth in males of L. squamosa . Further specimens are required to resolve the differences.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
|
Kingdom |
|
|
Phylum |
|
|
ParvPhylum |
Osteichthyes |
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lycenchelys squamosa Toyoshima, 1983
| Kawarada, Shumpei, Imamura, Hisashi, Narimatsu, Yoji & Shinohara, Gento 2020 |
Lycenchelys squamosa: Anderson, 1994: 113
| Nakabo, T. & Hirashima, Y. 2015: 217 |
| Hatooka, K. 2013: 1227 |
| Amaoka, K. & Nakaya, K. & Yabe, M. 2011: 318 |
| Balushkin, A. V. & Sheiko, B. A. & Fedorov, V. V. 2011: 983 |
| Shinohara, G. & Narimatsu, Y. & Hattori, T. & Ito, M. & Takata, Y. & Matsuura, K. 2009: 724 |
| Kitagawa, D. & Imamura, H. & Goto, T. & Ishito, Y. & Fujiwara, K. & Ueda, Y. 2008: 96 |
| Shinohara, G. & Anderson, M. E. 2007: 64 |
| Anderson, M. E. & Fedorov, V. V. 2004: 20 |
| Shiogaki, M. & Ishito, Y. & Nomura, Y. & Sugimoto, T. 2004: 71 |
| Hatooka, K. 2002: 1033 |
| Hatooka, K. 2000: 1033 |
| Imamura, H. 1997: 60 |
| Shinohara, G. & Endo, H. & Matsuura, K. 1996: 182 |
| Anderson, M. E. 1994: 113 |
Lycenchelys squamosus
| Zama, A. 2001: 86 |
| Imamura, H. 1998: 32 |
| Koyanagi, M. 1997: 538 |
| Amaoka, K. & Nakaya, K. & Yabe, M. 1995: 241 |
| Hatooka, K. 1993: 901 |
| Toyoshima, M. 1984: 293 |
| Toyoshima, M. 1983: 145 |