
Lycenchelys lenzeni Thiel, Knebelsberger & Eidus, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4370.1.3 |
|
publication LSID |
lsid:zoobank.org:pub:3A996552-91CD-4146-A610-7C737171FEAD |
|
DOI |
https://doi.org/10.5281/zenodo.5989755 |
|
persistent identifier |
https://treatment.plazi.org/id/01308795-FF9A-FFAD-35AA-33D1FE1DFD7A |
|
treatment provided by |
Plazi |
|
scientific name |
Lycenchelys lenzeni Thiel, Knebelsberger & Eidus |
| status |
sp. nov. |
Lycenchelys lenzeni Thiel, Knebelsberger & Eidus , sp. nov.
Simushir eelpout
( Figures 1–3 View FIGURE 1 View FIGURE 2 View FIGURE 3 ; Tables 1–3)
Holotype. ZMH 26255 , adult female, 296 mm SL, RV ‘Akademik M.A. Lavrentyev’, station 8-7, Bussol Strait, southwest of Simushir Island, Kuril Archipelago , Russia, 46°36.465' N, 151°34.615' E, Agassiz trawl, 2350 m depth, 24 July 2015 GoogleMaps .
*On epurals + on upper hypural plate + on lower hypural plate.
Paratypes. 6 specimens, same collection data as holotype; ZMH 26266, adult female, 296 mm SL, ZMH 26270, adult female, 285 mm SL, ZMH 26271, adult male, 263 mm SL, ZMH 26272, adult female, 242 mm SL, ZMH 26273, adult female, 226 mm SL, ZMH 26274, adult male, 182 mm SL.
Diagnosis. A species of Lycenchelys , distinguished from its congeners by the following combination of characters: vertebrae 26-28 + 100–102 = 126–130; interorbital and occipital pores absent; postorbital pores 3-4; suborbital pores 7 (rarely 6); preoperculo-mandibular pores 4 + 4; gill rakers 11–16; dorsal fin rays 118–122; anal fin rays 105-108; pelvic fin rays 2; middle and lower ray tips of pectoral fin very slightly exserted; lateral line double with mediolateral and ventral branches.
Description. Counts and measurements are provided in Table 1. Body relatively long ( Fig. 2 View FIGURE 2 ), its cross section almost round at nape and elliptical at abdominal and caudal regions.
Head elongate, ovoid, flat top, dorsal profile evenly declined from nape to relatively steeply sloping snout tip ( Fig. 3B View FIGURE 3 ). Eye ovoid, not entering dorsal profile of head. Interorbital space convex. Gill slit extending ventrally below the level of lower pectoral base. Opercular flap at upper margin of gill slit well developed. Mouth subterminal, upper jaw extending beyond anterior margin of pupil. Nostril tube not reaching upper lip when depressed forward. Snout rounded. Lower jaw with moderate labial lobe. Jaw teeth small, conical, in upper jaw in 2–3 rows anteriorly with outer ones larger, in lower jaw in 3–4 rows anteriorly, blending into single, short posterior row in both jaws. Vomerine teeth 10–17; palatine teeth 7–13 in 2 (rarely 1) irregular rows anteriorly, blending into single, posterior row. Gill rakers short and blunt; pseudobranch filaments relatively long.
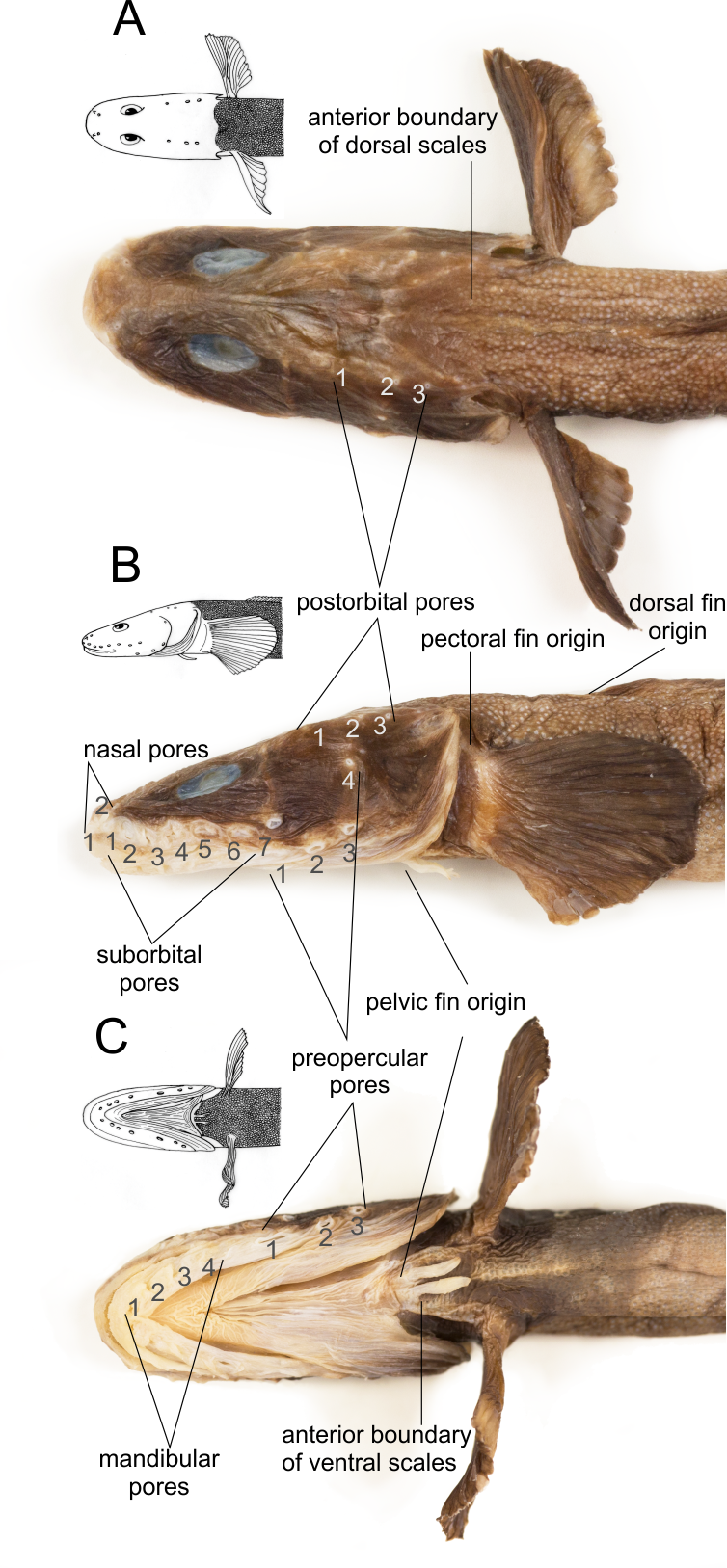
Head pores well developed ( Fig. 3 View FIGURE 3 ). Two nasal pores, anterior one set just in front of nostril tube, the other situated dorsoposteriorly. Seven (rarely six) suborbital pores arising from ventral ramus of bone chain under eye. Postorbital pores 3–4 arising from frontal-lateral extrascapular commissure. Eight preoperculomandibular pores (4 mandibular pores + 4 preopercular pores): 4 arising from dentary, one from anguloarticular and 3 from preopercle. Interorbital and occipital pores absent.
Body lateral line double, with mediolateral and ventral branches starting from above upper end of gill slit. Superficial neuromasts of both branches not detectable on tail, at the beginning of ventral branch rather dense, afterwards and over the whole course of mediolateral branch more widely separated.
Dorsal fin origin above middle of pectoral fin; lower tip of pterygiophore of first dorsal-fin ray is between tips of neural spines of sixth and seventh or seventh and eighth vertebrae. Pterygiophore of last dorsal ray is associated with fourth preural vertebra. Anal fin origin associated with ultimate or penultimate precaudal vertebra. Six or seven pterygiophores of anal fin inserted anteriorly to haemal spine of first caudal vertebrae; pterygiophore of last anal ray associated with second preural vertebra. Free pterygiophores absent. Pectoral fin origin at body midline, insertion on abdomen. Posterior margin of pectoral fin evenly rounded, middle and lower ray tips very slightly exserted, more so ventrally; ventralmost 5-7 rays thickened. Pelvic fin small, its length shorter than snout. Caudal fin with 2 epural, 4 upper and 4 lower hypural rays.
Scales absent on head, nape, thorax, pelvic fin, pectoral fin and mostly on pectoral fin base. Sometimes a few scales on lower part of pectoral fin basis. Scales present on body, abdomen, tail, on dorsal and anal fins, extending to about three-fourths of their height. Scales on dorsum before dorsal fin origin; extending anteriorly to a line connecting upper ends of gill openings, in larger specimens also a few scales somewhat in front of this line ( Fig. 3A View FIGURE 3 ). Belly scaled; extending anteriorly to basis of pelvic fin ( Fig. 3C View FIGURE 3 ).
Pyloric caeca not developed.
Color in alcohol. Head, pectoral fins and edges of dorsal and anal fins dark brown. Body brownish-grey, shifting from more brownish anteriorly to more greyish posteriorly. Eyes bluish-grey. Lips and ventral side of head whitish. Pelvic fins and area around anus white. Scales whitish, giving the darker anterior part of the body the appearance of being whitish spotted. Peritoneum dark brown. Branchial and oral cavity brownish.
Etymology. The species is named after Prof. Dr. Dieter Lenzen, President of Universität Hamburg and former President of Freie Universität Berlin, and one of the most cited and most influential educational theorists in Germany, in recognition of his outstanding support to the development of the research, educational and administrative facilities of the Center of Natural History (CeNak).
Distribution. Lycenchelys lenzeni is only known from the type locality.
DNA barcoding. For the four analysed paratypes ZMH 26271-74 full DNA barcodes (652 bp) were obtained and uploaded to GenBank (https://www.ncbi.nlm.nih.gov/genbank/) with the following accessions: MG650294 View Materials - MG650297 View Materials . Tissue samples and DNA isolates were stored in the DNA and tissue collection of the DZMB.
The four analysed paratypes share the same mitochondrial haplotype. The result of comparisons of the haplotype sequence of L. lenzeni with all published DNA barcodes on BOLD using the species identification engine are shown in Table 2. A match on species level could not be made supporting the species status of L. lenzeni . The closest match was with one specimen of Lycenchelys antarctica Regan, 1913 with a sequence similarity of 98.25%, followed by 5 specimens of Lycenchelys aratrirostris Andriashev & Permitin, 1968 (sequence similarities 97.95-97.96%) and one specimen of Lycenchelys jordani ( Evermann & Goldsborough, 1907) with a sequence similarity of 97.81%.
Comparative notes. The new species stems from the Western North Pacific, where 24 out of all known 62 Lycenchelys species occur ( Anderson & Balanov 2000, Anderson & Imamura 2002, Shinohara & Anderson 2007). Together with nine species found in the northeastern Pacific ( Froese & Pauly 2017), 33 Lycenchelys species are known from the North Pacific.
Amphiboreal species of Lycenchelys are not known. However, one North Atlantic species occurs also in the Arctic Ocean, but was not found in the North Pacific. Two other Lycenchelys species are endemic for the Arctic Ocean ( Priede 2017). The remaining species are endemic for the Atlantic, Indian and Southern Oceans and the South Pacific ( Priede 2017).
Although all specimens of L. lenzeni were caught in the bathyal zone (depth 2350 m), it cannot be considered as a typical “bathyal” species, since it has no interorbital and occipital pores and more precaudal vertebrae (26–28) than a typical “bathyal” species should have (less than 25) according to the definition of Andriashev (1955).
We have compared L. lenzeni with all 33 Lycenchelys species recognized for the North Pacific based on their morphological characters provided by Anderson (1995), Anderson & Balanov (2000), Anderson & Imamura (2002), Fedorov (1995), Fedorov & Andriashev (1993), Shinohara & Anderson (2007), Shinohara & Matsuura (1998) and Toyoshima (1985). Comparison of main characters is given in Table 3, where differences between the new species and the known species from the North Pacific are indicated in bold. Below are at least two different characters between the new species and each of all known 33 Lycenchelys species from the North Pacific are presented, further differences are shown in Table 3.
……continued on the next page The new species is different from fourteen species which occur in the North Pacific in having 126–130 total vertebrae and no interorbital pores ( Table 3). In comparison with the new species, the following nine out of the above mentioned fourteen species have lower numbers of total vertebrae (vertebrae counts for each species in brackets) and at least one interorbital pore: Lycenchelys alta Toyoshima, 1985 (88), Lycenchelys aurantiaca Shinohara & Matsuura, 1998 (85–88), Lycenchelys incisa (Garman, 1899) (106–110), Lycenchelys melanostomias Toyoshima, 1983 (121), Lycenchelys parini Fedorov, 1995 (125), Lycenchelys ratmanovi Andriashev, 1995 (110– 115), Lycenchelys squamosa Toyoshima, 1983 (91–94), Lycenchelys tohokuensis Anderson & Imamura, 2002 (116–119) and Lycenchelys vitiazi Andriashev, 1955 (114–116). The remaining five species have at least one interorbital pore and higher numbers of total vertebrae than L. lenzeni which are: Lycenchelys federovi Anderson & Balanov, 2000 (132–139), Lycenchelys hippopotamus Schmidt, 1950 (132-137), Lycenchelys maculata Toyoshima, 1985 (138-148), Lycenchelys makushok Fedorov & Andriashev, 1993 (136), and Lycenchelys rosea Toyoshima, 1985 (136–137).
Lycenchelys camchatica (Gilbert & Burke, 1992) (118–125) and Lycenchelys uschakovi Andriashev, 1958 (122) have lower vertebrae counts than the new species and only two postorbital pores, whereas L. lenzeni has 3–4 postorbital pores. Lycenchelys albeola Andriashev, 1958 (133) has also only two postorbital pores but higher vertebrae counts than the new species. In having lower vertebrae counts than the new species and three pelvic fin rays, Lycenchelys cicatrifer (Garman, 1899) (116–124), Lycenchelys folletti Anderson, 1995 (102-109), Lycenchelys porifer ( Gilbert, 1890) (113–120), Lycenchelys ryukyuensis Shinohara & Anderson, 2007 (118–124), and Lycenchelys scaurus (Garman, 1899) (96–110) differ from the new species which has only two pelvic-fin rays.
The remaining eleven North Pacific Lycenchelys species have overlapping vertebrae counts with L. lenzeni ( Table 3). These are the following species: Lycenchelys albomaculata Toyoshima, 1983 (123–130), Lycenchelys callista Anderson, 1995 (126–136), Lycenchelys crotalinus ( Gilbert, 1890) (121–131), Lycenchelys jordani ( Evermann & Goldsborough, 1907) (128–135), Lycenchelys micropora Andriashev, 1955 (123–135), Lycenchelys monstrosa Anderson, 1982 (124–132), Lycenchelys pearcyi Anderson, 1995 (128–134), Lycenchelys plicifera Andriashev, 1955 (122–126), Lycenchelys rassi Andriashev, 1955 (122–129), Lycenchelys remissaria Fedorov, 1995 (122–130), and Lycenchelys volki Andriashev, 1955 (126). However, L. albomaculata and L. rassi are different from the new species by having one interorbital pore (vs. no interorbital pore present in the new species). Additionally L. albomaculata is distinct from L. lenzeni by having 9 preoperculomandibular pores (vs. 8 preoperculomanibular pores in the new species) and L. rassi differs from the new species by having two occipital pores (vs. absent occipital pores in the new species). Compared to L. callista , L. crotalinus , L. pearcyi , L. plicifera , L. remissaria , and L. volki , the new species has only 6–7 suborbital pores (vs. 8 in L. remissaria and L. volki , 8–9 in L. plicifera , 8–10 in L. callista and L. crotalinus and 8–11 in L. pearcyi ). L. callista differs also from the new species in having two occipital pores (vs. absent occipital pores in the new species). L. pearcyi and L. monstrosa differ from the new species in having a lower number of postorbital pores (1–2 vs. 3–4 in the new species) and a higher number of preoperculomandibular pores (9 vs. 8 in the new species). Moreover, the new species differs from L. crotalinus and L. remissaria in having only two pelvic fin rays (vs. three pelvic fin rays in the above mentioned two species). In comparison with the new species, L. plicifera and L. volki have lower numbers of dorsal fin rays (110–115 and 113 vs. 118–122) and anal fin rays (99–102 and 101 vs. 105–108).
In having overlapping counts of vertebrae, dorsal, anal and pectoral fin rays, number of head pores and a double lateral line Lycenchelys lenzeni sp. nov. is morphologically most similar to L. micropora and L. jordani ( Table 3). However, L. micropora differs from the new species (two pelvic fin rays, pectoral fin origin at body midline) in having three pelvic fin rays and the pectoral fin origin below body midline. L. jordani differs from the new species in its higher number of pelvic fin rays (three vs. two). Furthermore, middle and lower ray tips of pectoral fin are very slightly exserted in Lycenchelys lenzeni sp. nov., whereas they are well exserted in L. jordani and L. micropora .
In addition to L. jordani from the eastern North Pacific, two species occurring in the Southern Ocean ( L. antarctica , L. aratrirostris ) showed the greatest sequence similarity with L. lenzeni . However, the two species from the Southern Ocean differ significantly from the new species on the basis of morphological characters. L. antarctica has a lower number of dorsal fin rays than L. lenzeni (102–116 vs. 118–122) and a much more retrograde origin of dorsal fin ( Anderson 1990). L. aratrirostris differs from the new species in having lower numbers of dorsal fin rays (97–107 vs. 118–122), anal fin rays (79–89 vs. 105–108), vertebrae (101–110 vs. 126–130) and a higher number of pelvic fin rays (three vs. two).
Comparative material
Lycenchelys antarctica : ZMH 9067, 1 specimen, 257 mm SL, off the South Shetland Islands, 60°37.10' S, 54°55.90' W, RV´Polarstern´, station 39/96, Agassiz trawl, 3213–3219 m depth, 26 November 1996. Lycenchelys aratrirostris : ZMH 9068, 5 specimens, 181–205 mm SL, off the South Shetland Islands, 62°18.90' S, 58°41.70' W, RV´Polarstern´, station 175/96, Agassiz trawl, 491–497 m depth, 24 December 1996. ZMH 121994, 1 specimen, 163 mm SL, Weddell Sea, 72°35.4' S, 18°06.8' W, RV´Polarstern´, station 273/8, Agassiz trawl, 562 m depth, 27 January 1985. ZMH 123481, 2 specimens, 200 and 241 mm SL, Weddell Sea, 69°58.9' S, 02°19.9' E, RV´Polarstern´, station 169, 140´, bottom trawl, 450–560 m depth, 19 February 1991. ZMH 123493, 1 specimen, 66 mm SL, Weddell Sea, 72°21.8' S, 16°52.1' W, RV´Polarstern´, station 158, 140´, bottom trawl, 539-633 m depth, 13 February 1991. Lycenchelys jordani : SIO 60-475, 5 from 9 specimens (1 cleared & stained), 146–360 mm SL, California, off Farallon Islands, otter trawl, ca. 1830 m depth, 10 November 1960. SIO 89-108, 1 from 2 specimens, 242 mm SL, California, San Clemente Basin, 32°36.1' N, 118°10.2' W, R. G. Sproul, 1500–1940 m depth, 12 September 1989.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |