
Klimakodesmus bilobocaudatus, Sudhikumar, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4980.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:C55A445F-76EC-460A-B259-98AF744ECFFF |
|
DOI |
https://doi.org/10.5281/zenodo.4986346 |
|
persistent identifier |
https://treatment.plazi.org/id/024F8783-5967-7F30-C5FD-7A94FAB622D6 |
|
treatment provided by |
Plazi |
|
scientific name |
Klimakodesmus bilobocaudatus |
| status |
sp. nov. |
Klimakodesmus bilobocaudatus sp. nov.
Figs 1–15 View FIGURES 1–4 View FIGURES 5–7 View FIGURES 8–15
Material examined. Adult male holotype (CATE-5302), 2 adult male and 6 subadult female paratypes (CATE- 5302B–CATE-5302I), GoogleMaps 1 adult male and 2 subadult female paratypes ( ZMUM), India, Kerala state, Kannur district, N11°58’2.5”, E75°17’46.6”, 5 m a.s.l., 27/01/2021, M.D. Aswathy leg. GoogleMaps
Name. To emphasize a caudally relatively deeply bilobed dorsal keel ( PM) on ring 19 in the new species; adjective.
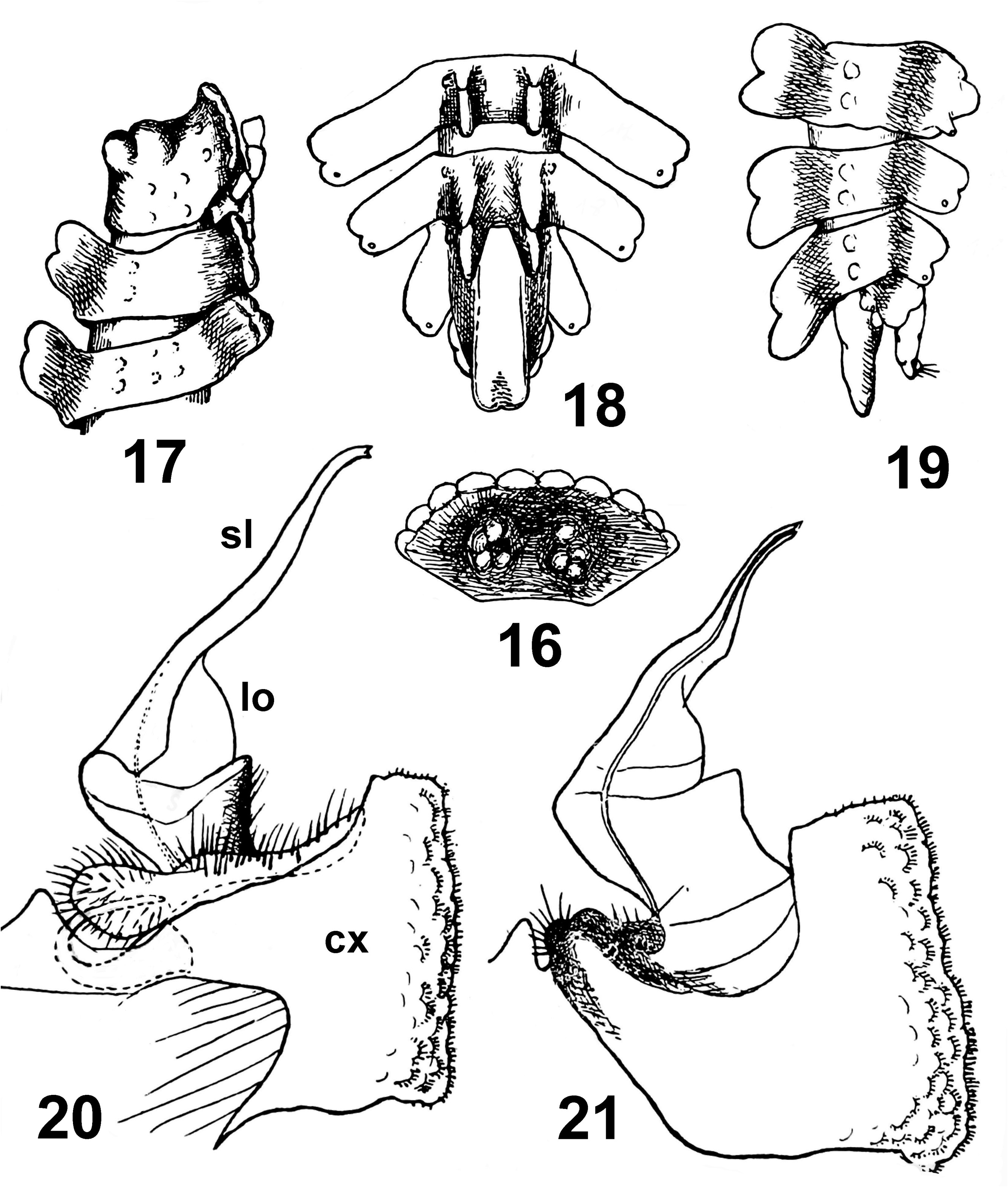
Diagnosis. The new species differs clearly from K. gravelyi , the only other congener to be accepted ( Carl 1932), by the more regularly tuberculate collum (cf. Figs 5 View FIGURES 5–7 and 16 View FIGURES 16–21 ), the lower PM and DL which are mostly inclined caudad (vs. inclined forward until ring 16, cf. Figs 7 View FIGURES 5–7 , 8 View FIGURES 8–15 and 17 View FIGURES 16–21 ), the caudally more deeply bilobed dorsal keel (PM) on ring 19 (vs. indistinctly bilobulate, cf. Figs 9, 10 View FIGURES 8–15 and 18 View FIGURES 16–21 ), the laterally bi- (rings 3 and 4) or trilobate (remaining postcollum rings) paraterga (vs. mostly bilobate, trilobate only on ring 2, cf. Figs 5 View FIGURES 5–7 , 8, 9 View FIGURES 8–15 and 17–19 View FIGURES 16–21 ), and the gonopods showing subspherical coxites (vs. squarish) and subcontiguous, apically subunciform and crossing each other (vs. divergent and suberect) telopodites, each of the latter additionally reinforced with a mesobasally thickened wall (vs. a laterobasal lobe, cf. Figs 12–15 View FIGURES 8–15 and 20 & 21 View FIGURES 16–21 ).
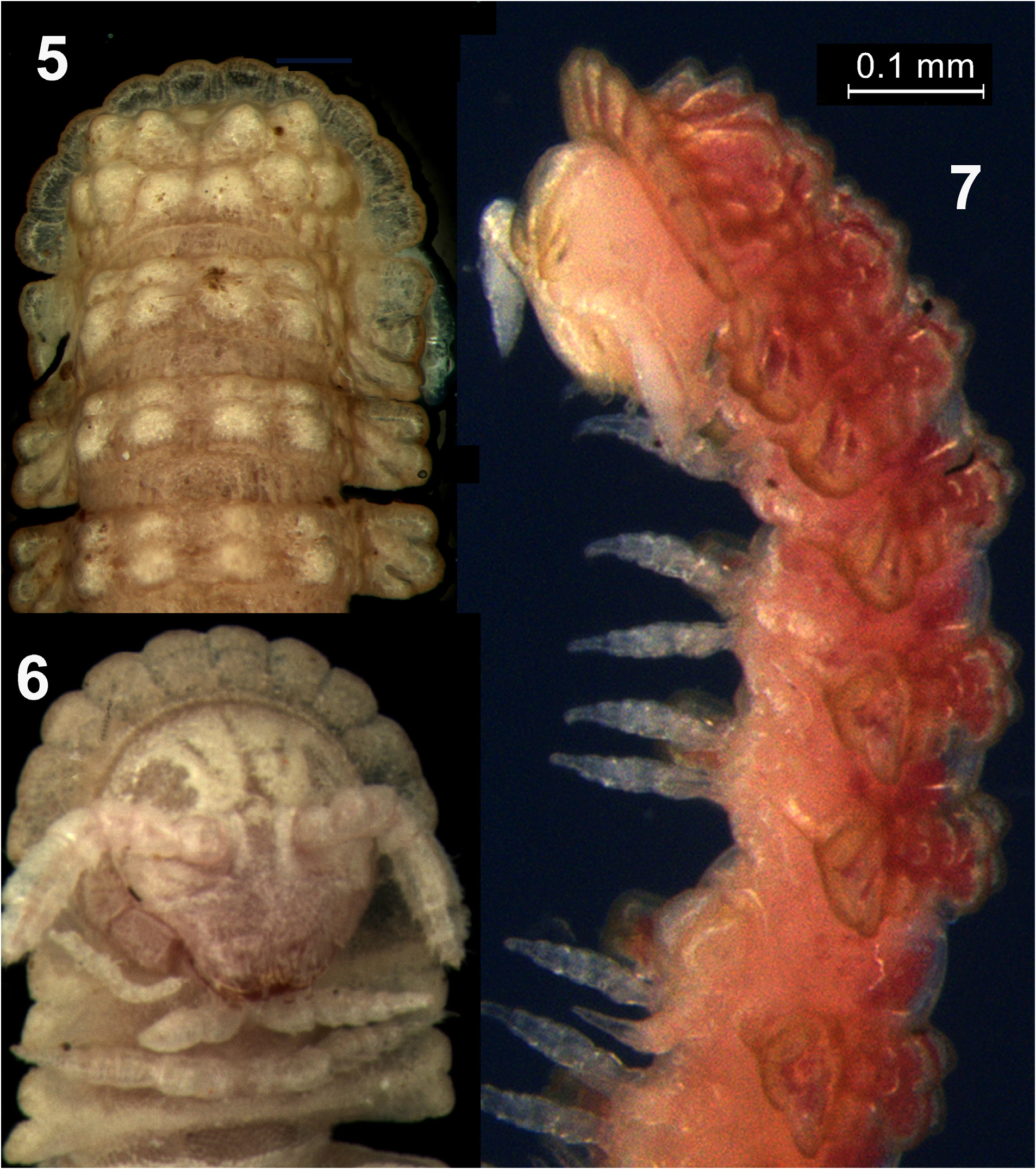
Description. Length ca. 3.68 mm, width of midbody segments 0.10 and 0.14 mm on pro- and metazonae, respectively (holotype). Length of adult male paratypes ca. 3.0– 3.8 mm, that of subadult female paratypes 3.2–3.5 mm. Width of adult male paratypes ca. 0.40–0.45 mm, that of subadult female paratypes 0.44–0.48 mm. Live colouration generally pink; antennae, legs and venter translucent, lighter than dorsum ( Figs 1 View FIGURES 1–4 , 7 View FIGURES 5–7 & 9 View FIGURES 8–15 ). Colouration in alcohol after 2 months of preservation pale brown to pinkish brown, legs and antennae faded to translucent ( Figs 2–6 View FIGURES 1–4 View FIGURES 5–7 , 8 & 10 View FIGURES 8–15 ). No earth crust visible on body.
Most characters as in the generic description above, except as follows.
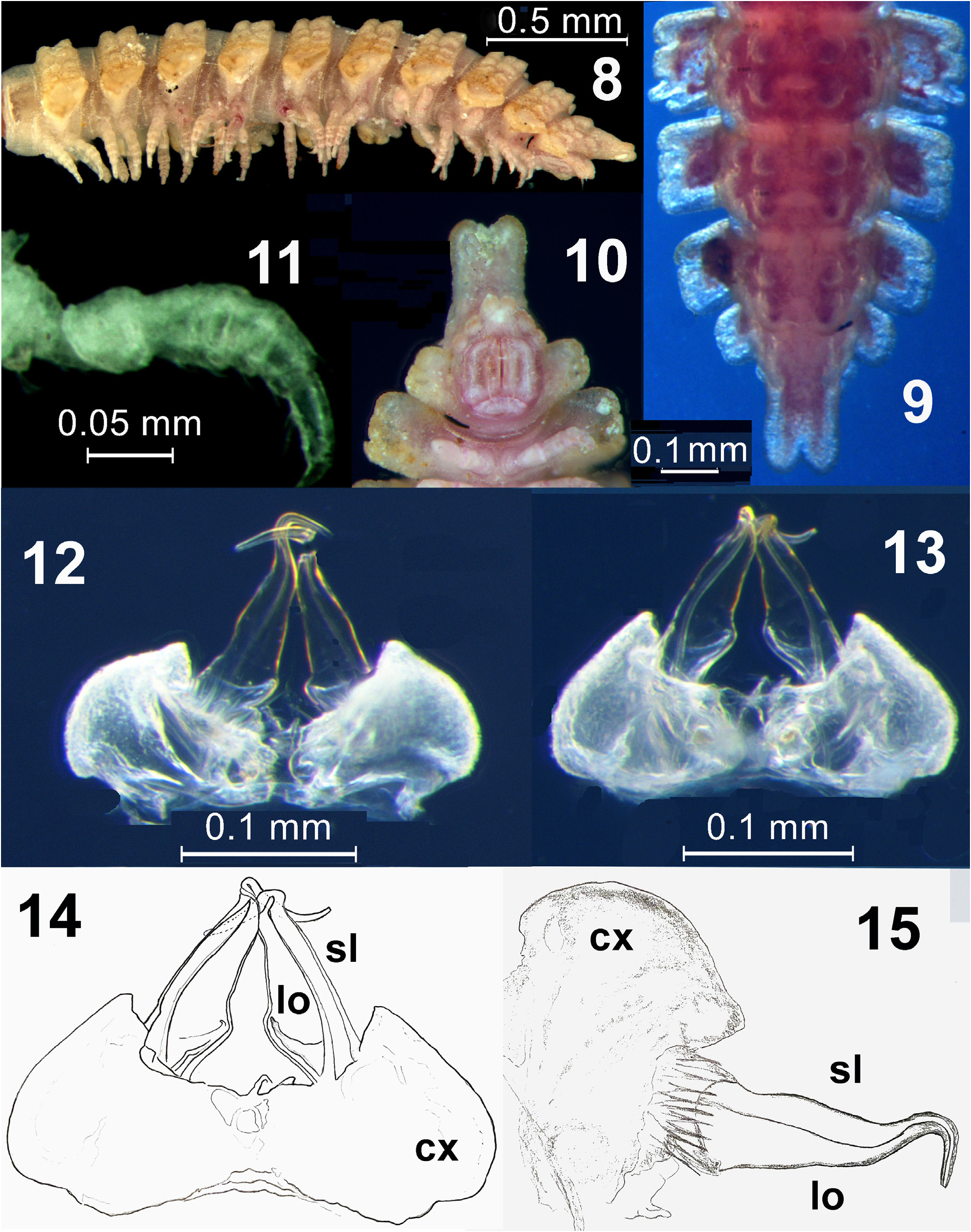
Body with 20 segments (likely in both sexes, see below) ( Figs 2 & 4 View FIGURES 1–4 ). In width, head << collum <segment 2=3 <4–6 <7–9 <10–15, body gradually tapering towards telson thereafter. Head circular, labral region moderately setose, epicranial suture rather deep, vertex microgranulate ( Fig. 6 View FIGURES 5–7 ). Interantennal isthmus nearly as wide as diameter of antennal socket. Antennae short, weakly clavate, in situ reaching body segment/ring 3 when stretched ventrolaterally. In length, antennomere 7<3<1<6<4<2<<5; antennomeres 5–7 each with a more or less compact apicodorsal group of bacilliform sensilla ( Figs 6 & 7 View FIGURES 5–7 ). Collum flabellate, completely covering the head from above; anterior margin with 5+5 distinct, equal, moderately incised and rounded lobulations; central part domed, with 2+2 and 3+3 prominent and rounded tubercles arranged in two transverse rows ( Figs 2–7 View FIGURES 1–4 View FIGURES 5–7 ). Paraterga set low (at about half of midbody height), subhorizontal to faintly declivous. Dorsum strongly arched, its outline smoothly extending onto paraterga ( Figs 3 View FIGURES 1–4 & 7 View FIGURES 5–7 ). Prozonae, as well as deep and thin strictures between pro- and metazonae finely alveolate. Postcollum metaterga with strongly differentiated dorsal tuberculations: 2+2 longitudinal crests/rows of mostly 2+2 fused tubercles, those before ring 19 slightly inclined caudad, but PM on ring 19 abruptly modified into a mid-dorsal, strongly elongate, conspicuous and caudally clearly bilobate ridge overhanging and concealing a short epiproct ( Figs 2–4 View FIGURES 1–4 , 8–10 View FIGURES 8–15 ). Paraterga broad, thin, rather irregularly tuberculate near bases and clearly lobulated laterally, with three rounded and moderately incised lobulations on rings 2–19, regardless of cylindrical porosteles located between the 2 nd and 3 rd lobulations on rings 5, 7, 9, 10, 12, 13, 15 and 16 ( Figs 2–10 View FIGURES 1–4 View FIGURES 5–7 View FIGURES 8–15 ); only paraterga 3 and 4 with last lobulation in a clearly caudolateral position ( Figs 5–7 View FIGURES 5–7 ). Epiproct ( Fig. 10 View FIGURES 8–15 ) finger-shaped, tip bent down and divided into two round bulbs. Hypoproct ( Fig. 10 View FIGURES 8–15 ) semi-circular, with 1+1 strong setae borne on minute, caudal, paramedian knobs.
Legs with neither adenostyles nor tarsal brushes ( Fig. 11 View FIGURES 8–15 ). In length, tarsi> femora> prefemora> tibiae> coxae> postfemora; claws simple, slightly curved ventrally.
Gonopods ( Figs 12–15 View FIGURES 8–15 ) relatively simple, placed inside a transversely oblong-oval gonopodal aperture, the latter with thin and slightly elevated caudal and lateral margins; in situ, both gonopods strongly exposed, held mostly parallel to one another, with only tips crossing each other. Coxites (cx) voluminous, but moderately enlarged, gonocoel thus being relatively shallow; cannulae simple. Telopodites directed caudally, each unipartite and subtriangular, long and slender, gradually attenuating distad and acuminate apically, each represented solely by a solenomere (sl) and supporting a seminal groove running along lateral margin to the end of a laterally directed and unciform apical part, and additionally reinforced basally through a distinctly thickened caudomesal wall of a prominent, membranous, mesal lobe (lo); prefemorite as usual, relatively short, densely and strongly setose.
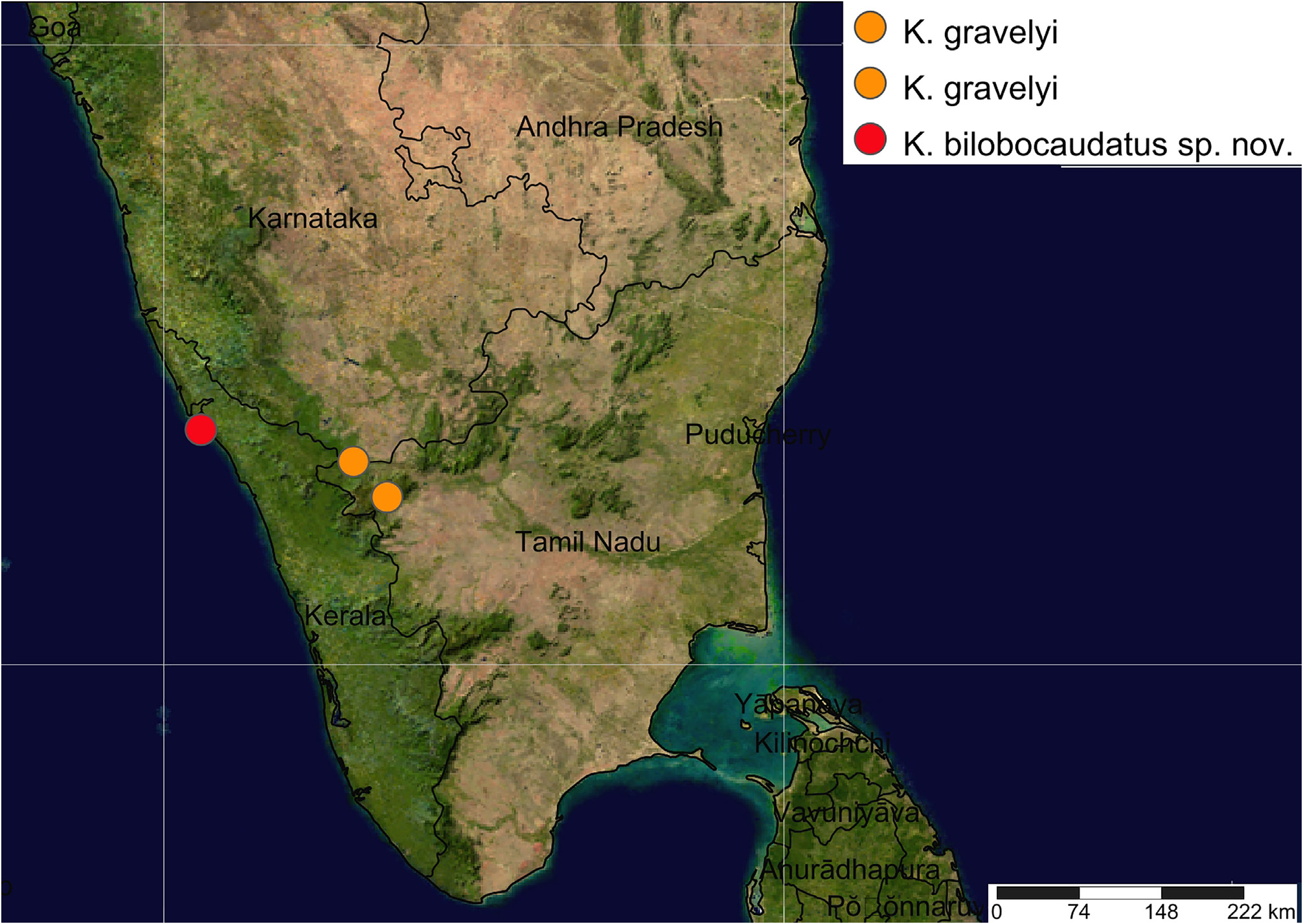
Ecological notes. This new species was found under rotting wood near a lowland swamp overgrown with Myristica forest in the Kannur district, northern Kerala, India. The habitat area of approximately 20 ha is a sacred grove protected by local people. In contrast, K. gravelyi came from high-altitude areas (1000–1600 m a.s.l.) in the state of Tamil Nadu ( Carl 1932), Western Ghats, southern India ( Fig. 22 View FIGURE 22 ).
It seems noteworthy that all females collected at the type locality in January appear to be subadults, each with 19 body segments. This has been proven by dissecting the vulvae which have always been found underdeveloped (amorphous ovoid clods devoid of setae), and an anterior, ventral, transverse, marginal ridge on ring 3 missing. This agrees well with the common wisdom that only adult males in Polydesmida , but never adult females, can be composed of a lesser number of body rings. Usually, in the course of teloanamorphosis characteristic of the order Polydesmida , both the males and females of Polydesmida show the same number of body rings, either 20 or 19, more rarely 18, exceptionally up to 40 ( Shear et al. 2016). When there is a difference in polydesmidan body segment counts, the adult males typically have one segment less and nearly always remain stable per sex ( Enghoff et al. 2015). Only exceptionally, as is the case of Ammodesmus granum Cook, 1896 and A. congoensis VandenSpiegel & Golovatch, 2015 (Ammodesmidae) , from western or central Africa, respectively, can both sexes vary in the number (18 or 19) of body rings ( VandenSpiegel & Golovatch 2012, 2015).
In Klimakodesmus species , however, the situation is ordinary, as both sexes (must) equally have 20 body segments. The absence of adult females in the type series seems best to be accounted for by seasonal factors alone.

| ZMUM |
Zoological Museum, University of Amoy |
| PM |
Pratt Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |