
Trachysalambria longipes
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4150.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:323C3A73-8564-470D-94B0-4A71DAE9E940 |
|
DOI |
https://doi.org/10.5281/zenodo.5627600 |
|
persistent identifier |
https://treatment.plazi.org/id/03818796-FFCB-F928-C0C9-824061A9FA70 |
|
treatment provided by |
Plazi |
|
scientific name |
Trachysalambria longipes |
| status |
|
Trachysalambria longipes View in CoL (Paul’son, 1875)
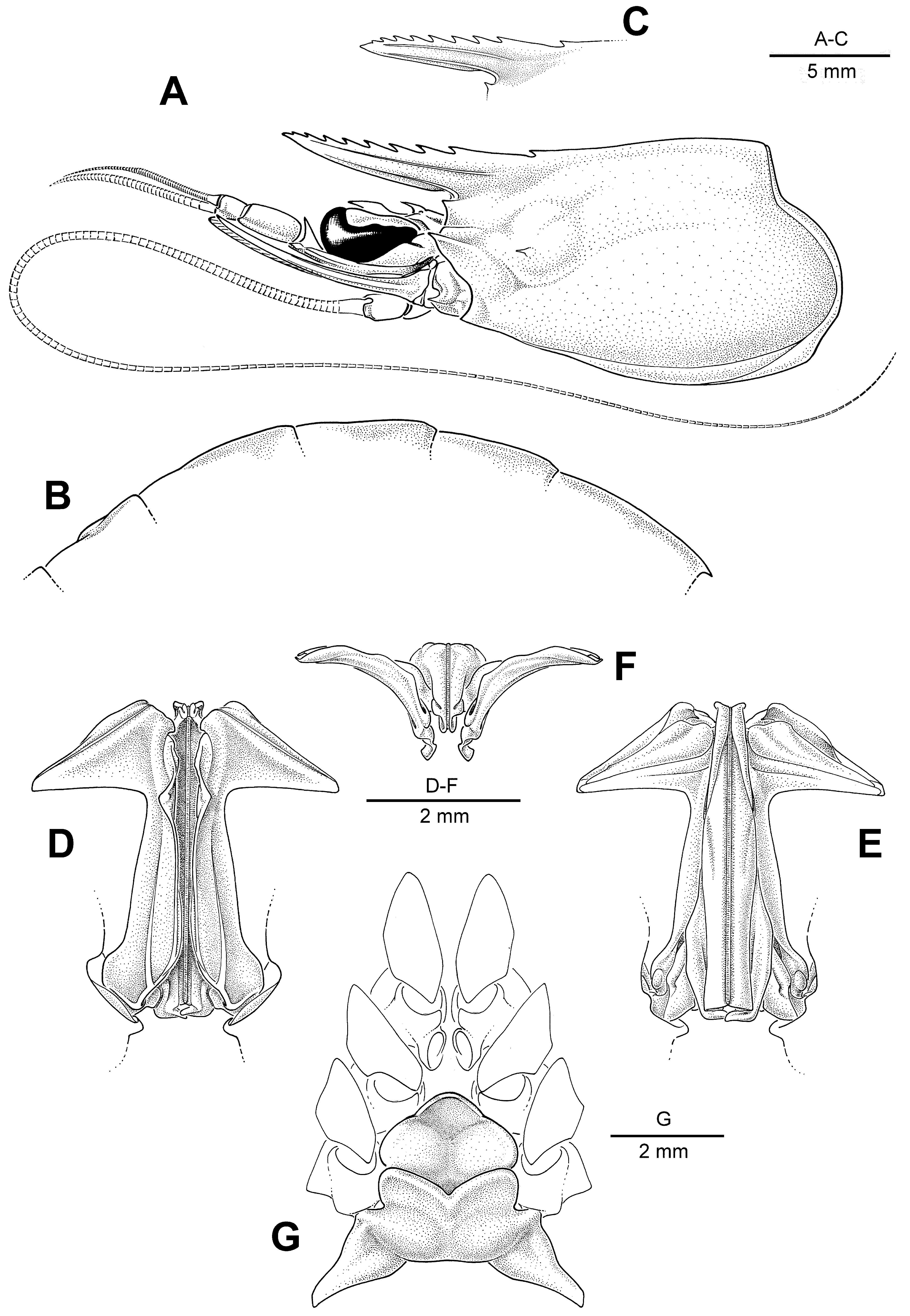
( Fig. 2 View FIGURE 2 )
Penaeus longipes Paul’son, 1875: 125, table 19-fig. 1, 1a [type locality: Red Sea].
Trachypenaeus villaluzi Muthu & Motoh, 1979: 58 View in CoL , text-figs. 1–2 [type locality: the Philippines]; Motoh & Buri, 1984: 89, figs. 61–62; Chan, 1998: 951, unnumbered figs.
Trachypenaeus curvirostris View in CoL .— Holthuis, 1980: 53 (in part). [not Stimpson, 1860]
[Not] Trachypenaeus longipes .— Motoh & Buri, 1984: 84, figs. 57–58; Liu & Zhong, 1988: 187, fig. 116, pl. 4-1; Hayashi, 1986: 77, fig. 37; 1992: 144, figs. 75b, 76c, 77c, 78c; Chan, 1998: 950, unnumbered figs. [= T. dentata sp. nov.]
Trachysalambria longipes View in CoL .— Pèrez Farfante & Kensley, 1997: 149 (in part); De Gave & Fransen, 2011: 228. (in part).
[Not] Trachysalambria longipes View in CoL .— Pèrez Farfante & Kensley, 1997: 149 (in part); Sakaji & Hayashi, 2003: 159, fig. 6; De Grave & Fransen, 2011: 228. (in part). [= T. dentata sp. nov.]
Trachysalambria villaluzi View in CoL .— Pèrez Farfante & Kensley, 1997: 149; De Grave & Fransen, 2011: 228.
Type material. Neotype: Seychelles, REVES II, stn CH 0 6, 4°57.8’S, 56°12.0’E, 40 m, 0 4.09.1980, female cl 25.5 mm ( MNHN IU- 2014-7043 ) GoogleMaps .
Other material examined. Philippines. Panay Is., Tigbauan, otter trawl, 26.12.1978, 5 males cl 9.3–10.5 mm (1 without cephalothorax), 7 females cl 12.0–15.0 mm (MNHN IU-2014-7045, ex SEAFDEC Aquaculture Department Museum, D288, paratypes of T. villaluzi ).
Indonesia. CORINDON II, stn CH 201, 01°11’S, 117°06’E, 21 m, 30.10.1980, 1 female cl 18.0 mm ( MNHN IU- 2014-7046 ). GoogleMaps
Fiji. SUVA 2 View Materials : stn CP 46, 17°52.5’S, 177°15.5’E, 25 m, 19.10.1998, 1 male cl 8.2 mm ( MNHN IU- 2014- 7050 ) GoogleMaps ; stn CP 47, 17°53.5’S, 177°13.6’E, 25 m, 19.10.1998, 7 males cl 7.0–9.0 mm, 1 female cl 9.5 mm (MNHN IU-2014-7049); stn CP 57, 17°43.4’S, 177°22.8’E, 17 m, 20.10.1998, 1 male cl 7.0 mm (MNHN IU-2014-7048), 1 female cl 9.2 mm (MNHN IU-2014-7047).
Red Sea. Hamar Is., Kamaran , no date, 1 female cl 15.5 mm ( MNHN IU- 2014-7051 ).
Seychelles. REVES II, stn CH 0 6, 4°57.8’S, 56°12.0’E, 40 m, 0 4.09.1980, 2 females cl 21.5 and 22.2 mm ( MNHN IU- 2014-7044 ). GoogleMaps
La Réunion. “ Marion-Dufresne ”, MD/32: stn CP 42, 21°20.7’S, 55°26.9’E, 74–77 m, 18.08.1982, 1 female cl 18.5 mm ( MNHN IU- 2014-7057 ) GoogleMaps ; stn CP 125, 20°52.5’S, 55°36.9’E, 45 m, 0 2.09.1982, 3 females cl 17.0–19.0 mm (MNHN IU-2014-7055); stn CP 127, 20°52.0’S, 55°37.1’E, 92 m, 0 2.09.1982, 1 female cl 17.0 mm (MNHN IU-2014-7056); stn CP 174, 20°51.8’S, 55°36.5’E, 78–85 m, 0 8.09.1982, 3 females cl 12.7–19.0 mm (MNHN IU- 2014-7058).
Madagascar. “ Vauban ”, Antongil Bay, trawl, 15°38.8’S, 49°42.4’E, 20 m, 0 2.04.1973, 2 males cl 13.0 and 16.2 mm, 16 females cl 13.6-20.1 mm ( MNHN IU- 2014-7052 ), 13 males cl 11.1–16.7 mm, 20 females cl 13.6– 21.2 mm ( MNHN IU- 2014-7053 ), 5 males cl 12.5–17.0 mm, 1 female cl 18.5 mm ( MNHN IU- 2014-7054 ). GoogleMaps
Description. Entire body densely pubescent. Rostrum with 7–10 (usually 8 or 9, excluding epigastric tooth) teeth along entire dorsal border; slightly to distinctly curved upwards in females, with ventral border more or less convex, tip straight or very slightly recurved downwards, with ventral margin more or less straight, tips of rostral teeth aligned in concave configuration; in males, rostrum straight to slightly curving upwards or downwards, ventral border convex to straight, tip more or less straight, tips of rostral teeth aligned in a straight or low crest configuration; more or less extending to tip of second segment of antennular peduncle (generally shorter in males and small females); postrostral carina from blunt to sharp, generally extending to near posterior carapace though sometimes rather indistinct in posterior half. Only pereiopods II and III bearing epipods. Pereiopod I generally bearing small ischial spine, which sometimes very minute and occasionally even completely absent. Pereiopod IV in females with coxa not particularly expanded medially. Pereiopod V more or less extending to tip of scaphocerite. Abdomen with dorsal carinae distinct on somites II to VI; that on somite II short; that on somite III distinct at posterior 2/3 of somite and rudimentary or absent at anterior 1/3 of somite; ridges on somites IV and V posteriorly incised and not terminating in distinct spine. Telson with strong but blunt dorsolateral carinae, bearing 3 or 4 pairs of movable lateral spines. Male petasma with lateral margins of stem distinctly converging distally; distolateral lobes with ventral margins more or less straight, tips of dorsal and ventral flaps coinciding, forming sharp angle. Female thelycum with anterior plate semi-triangular but with lateral parts somewhat protruding; anterior margin of posterior plate distinctly concave, with median notch.
Coloration. The coloration of the Philippines material is described as pale reddish brown. Antennal flagella milky or pinkish white. Posterior margins of abdominal somites with narrow transverse light brownish bands. Uropods brownish red to reddish with white margins ( Muthu & Motoh 1979; Motoh & Buri 1984).
Distribution. Indo-West Pacific and known with certainty from Madagascar, La Réunion, Seychelles, Red Sea, Indonesia, the Philippines and Fiji; at depths of 7– 92 m.
Remarks. The present species, generally known under the name T. villaluzi ( Muthu & Motoh, 1979) , is unique in the genus by lacking epipod on the pereiopod I but having epipods on the pereiopods II and III. Trachysalambria villaluzi was thought to be restricted to the Philippines ( Motoh & Buri 1984; Pèrez Farfante & Kensley 1997) but it is actually widely distributed in the Indo-West Pacific and with low genetic divergence (0.6% sequence divergence in 12S rRNA gene between materials from Seychelles and Fiji, Table 2). On the other hand, T. villaluzi is in fact very similar in general morphology to T. aspera , which also has a similar geographical distribution in the Indo- West Pacific. Although Muthu & Motoh (1979) and Motoh & Buri (1984) argued that the petasma and thelycum are different between T. villaluzi and T. aspera , the genitalia structures of the present species actually have the general shape of the genus ( Fig. 2 View FIGURE 2 D–G) and with large variations.
As discussed in the “Remarks” under P. dentata sp. nov., the “ T. longipes ” material previously reported from the Western Pacific and the South China Sea is actually not Paul’son’s (1875) species from the Red Sea. The exact identity of Penaeus longipes Paul’son, 1875 is uncertain as the type cannot be located now (M. Türkay pers. comm.; also see Sakaji & Hayashi 2003). Paul’son’s (1875) original description and thelycum illustration (Paul’son 1875: table 19-fig. 1a) clearly show that his specimen is a Trachysalambria . The original description given for the single type of T. longipes is rather brief but contains the following key characters used in this study for distinguishing the species of Trachysalambri a. The type is a mature female with a slightly curved rostrum. The postrostral carina extends to the posterior carapace. The pereiopod V extends to the distal end of scaphocerite. Dorsal carinae are distinct on the abdominal somites III to VI but only that on the somite VI bearing a posterior spine. Thelycum has the anterior plate semi-triangular and the posterior plate anteriorly with a median cleft.
Of the species in this genus, only T. villaluzi and T. aspera fit well with Paul’son’s (1875) original description and both are confirmed to occur in the Red Sea. Although T. palaestinensis is the commonest species in the Red Sea, it clearly differs from Paul’son’s (1875) species in the postrostral carina only extending to the posterior 2/3 of the carapace and the pereiopod V generally reaching far behind the tip of scaphocerite. The other species of Trachysalambria known with certainty in the western Indian Ocean is only T. parvispina sp. nov. However, as in T. dentata sp. nov., the dorsal carinae of the abdominal somites IV and V terminate in spines in T. parvispina sp. nov. Trachysalambria villaluzi and T. aspera can only be satisfactorily separated by the pereiopod I having an epipod in the latter but not in the former. Paul’son’s (1875: fig. 1) original figure shows a well-developed epipod on the basal parts of one of the chelipeds. However, the caption of this figure only states “One of the chelipeds” and there is no mention in the original description about the epipods on the chelipeds. There is a strong basial spine in Paul’son’s (1875: fig. 1) material but both pereiopods I and II have strong basial spines in all the members of Trachysalambria (as pereiopod III lacks basial spine in Trachysalambria , therefore, T. longipes must not be T. malaiana which lacks epipod on both pereiopods I and II). Since it cannot be known now if Paul’son’s (1875) fig. 1 is on pereiopod I or II, it is impossible to determine if T. longipes (Paul’son, 1875) belongs to the same species as T. villaluzi or T. aspera . The name T. longipes is widely but incorrectly used in recent literature, and many distinguishing characters for the members of Trachysalambria are now re-defined. Therefore, it is necessary to fix the name T. longipes by selecting a neotype. Between T. villaluzi and T. aspera , T. aspera is characterized by the female types having a very straight rostrum (see Alcock 1906: pl. 9-fig. 28), which is somewhat different from the original description of T. longipes . Moreover, specimens of these two species in the Indian Ocean often differ in the posterior half of the postrostral carina being distinct in T villaluzi but often indistinct in T. aspera . Since Paul’son’s (1875) type has a distinct postrostral carina extending to posterior carapace, a specimen of T. villaluzi is here selected as the neotype of T. longip es. By doing so, T. villaluzi effectively becomes a junior synonym of T. longip es. The type locality of T. longipes is from the Red Sea. However, the only Red Sea specimen of the present species available in this study is an old and damaged female (not in good condition with pereiopods mostly missing and tip of rostrum also missing). Other available material of this species nearest to the Red Sea is from Seychelles. Thus, a Seychelles female with genetic data ( Table 1 View TABLE 1 ; MNHN IU-2014-7043) is selected as the neotype of T. longipes .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Trachysalambria longipes
| Chan, Tin-Yam, Cleva, Régis & Chu, Ka Hou 2016 |
Trachysalambria longipes
| Gave 2011: 228 |
| Farfante 1997: 149 |
Trachysalambria villaluzi
| De 2011: 228 |
| Farfante 1997: 149 |
Trachypenaeus curvirostris
| Holthuis 1980: 53 |
Trachypenaeus villaluzi
| Chan 1998: 951 |
| Motoh 1984: 89 |
| Muthu 1979: 58 |