
Diploneis limosa, Jovanovska & Wilson & Hamilton & Stone, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.593.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.7875231 |
|
persistent identifier |
https://treatment.plazi.org/id/038487E2-FF86-261E-BCF1-FF09B92472B4 |
|
treatment provided by |
Plazi |
|
scientific name |
Diploneis limosa |
| status |
sp. nov. |
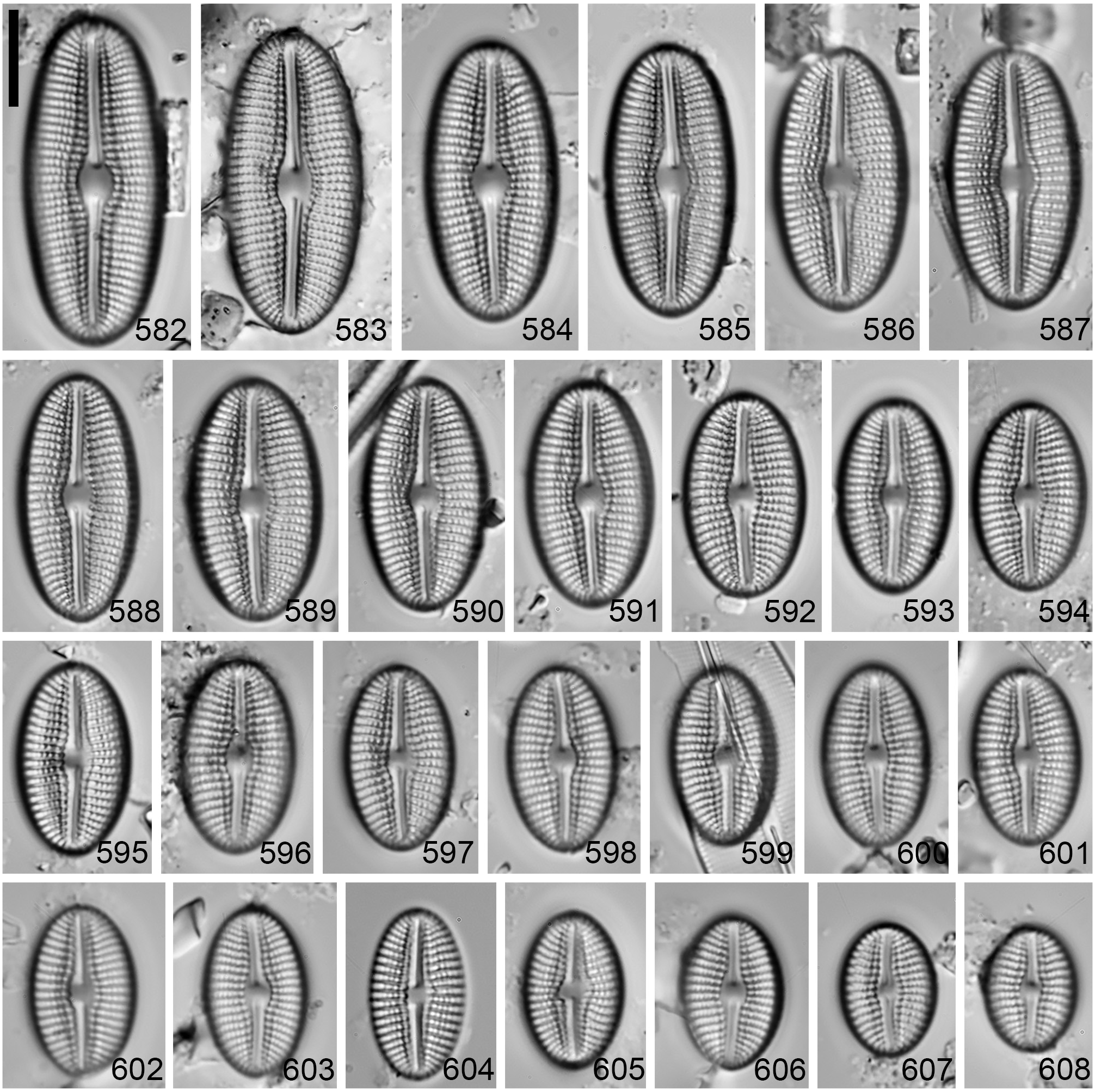
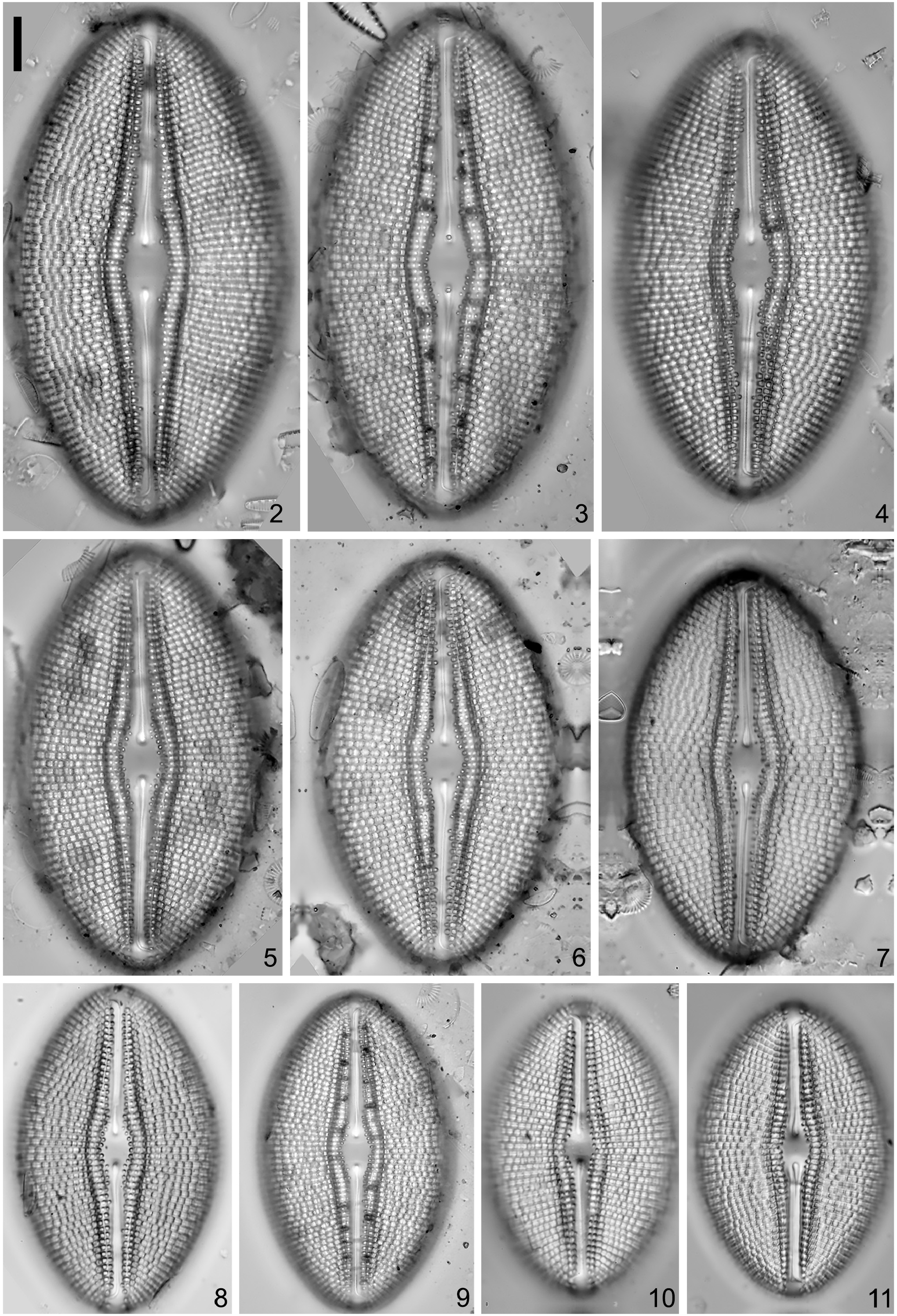
Diploneis limosa sp. nov. (LM Figs 582–608 View FIGURES 582–608 , SEM Figs 609–618 View FIGURES 609–613 View FIGURES 614–618 )
Valves are weakly asymmetric, linear-elliptic with parallel to weakly convex margins and round apices ( Figs 582– 609 View FIGURES 582–608 View FIGURES 609–613 ). Valves become elliptic-circular with smaller cell size. Valve length is 12.5–33.5 μm and width is 9.5–14 μm. The axial area is very narrow, lanceolate, slightly expanding into a longitudinally elongate central area ( Figs 584 View FIGURES 582–608 , 609, 610 View FIGURES 609–613 ), 2.2–3.9 μm wide. Externally, the canal is linear to lanceolate, slightly expanded in the middle of the valve with one row of cribrate (<20 poroids) areolae that widens into two along the raphe branches and then narrows into one at the valve apices ( Figs 582–585 View FIGURES 582–608 , 609, 611 View FIGURES 609–613 ). Internally, a thick non-porous slightly raised silica plate encloses the longitudinal canal ( Fig. 614 View FIGURES 614–618 ). Externally, the raphe is filiform, curved; the proximal ends are deflected to one side and positioned within an expanded teardrop depression ( Figs 609, 610, 611 View FIGURES 609–613 ). The distal raphe ends are unilaterally bent to the same side as the proximal ends and terminate on the upper mantle at the apex ( Figs 609, 611, 613 View FIGURES 609–613 ). Internally, the raphe is curved with simple proximal and distal ends that are slightly elevated within a depression formed by the longitudinal canal ( Figs 614, 616–618 View FIGURES 614–618 ). The striae are parallel at mid-valve changing to radiate towards the apices, 12–14 in 10 μm. Striae are uniseriate becoming biseriate towards the valve margins ( Fig. 612 View FIGURES 609–613 ). The striae are composed of round recessed areolae covered externally with fine pored cribra (15–40 poroids), 15–20 in 10 μm. The areolae increase in size towards the valve margins ( Figs 609, 611, 612 View FIGURES 609–613 ). Internally, the alveoli open via a single elongated opening covered with a thin silica layer ( Fig. 615 View FIGURES 614–618 ).
Type:— REPUBLIC OF ZAMBIA, Lake Mweru Wantipa , at 934 m elevation; floating plant roots, 0.1 m water depth, 8°33’33.2” S 29°56’15.4” E, E. Jovanovska & Z. Levkov, 3 rd October 2021 (holotype designated here, circled specimen BM-108974! = Fig. 583 View FIGURES 582–608 , GoogleMaps isotypes ANSP-GC17203 !, CANA-129327!). Type material CANA-129327. Registration: http://phycobank.org/103712 GoogleMaps
Pictures of the isolated specimen:— LM micrograph on 1000× magnification ( Fig. S3r View FIGURES 2–11 ).
Sequence data:— Plastid gene rbc L sequence (GenBank accession: OQ 660298).
Etymology:— The specific epithet ‘ limosa ’ refers to Lake Mweru Wantipa or ‘muddy lake’ where the species was originally discovered.
Ecology and distribution:— This species has only been observed in Lake Mweru-Wantipa, where the pH is neutral (7.3), the conductivity is high (1114 μS̔ cm-1), and the transparency of the water is very low. This was the only Diploneis species we found in the samples from the lake, and it was extremely abundant.
Main differential characters:— Valve size and shape, striae pattern, striae density, and poroids up to 40 per areola.
Similar species:— Diploneis fontium and D. calcifuga .
Phylogenetic relationships
ML and BI phylogenetic analyses on the single rbc L and concatenated rbc L+18S datasets revealed a nearly congruent topological pattern ( Figs 620 View FIGURE 620 , S 5 View FIGURES 2–11 ). The concatenated dataset yielded higher support for basal nodes than those inferred from the single locus dataset in both phylogenies. The two phylogenetic analyses supported monophyly of East African taxa with two distinct and well-supported clades, of which one clade included species from Lake Tanganyika (BS = 94 %, BPP = 1 for rbc L; BS = 93 %, BPP = 0.99 for rbc L+18S) and Diploneis limosa from Lake Mweru Wantipa as sister with 95 % BS support for both rbc L and rbc L+18S ( Fig. 620 View FIGURE 620 ). Within the Tanganyika clade, two entities remained unidentified, listed herein as Diploneis sp. 1 and Diploneis sp. 2 (see Table S1 View TABLE 1 ). The second clade consists of two species from Mulungushi River (BS = 73 %, BPP = 0.99 for rbc L; BS = 81 %; BPP = 0.99 for rbc L+18S). Support values within the Tanganyika clade and clade relationships are low in both analyses, but especially so in the ML analysis (BS <69 %). Therefore, neither BI nor ML support the morphological grouping of species based on valve outline and striae pattern, possibly due to low genetic coverage or genetic variation in both, rbc L and 18S, markers. All lineages differ by 0.05 nucleotide substitutions per site (overall mean p -distance) when assuming uniform rates for rbc L (Table S4).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |