
Physalia, Lamarck, 1801
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4669.1.1 |
|
publication LSID |
urn:lsid:zoobank.org:pub:6E2F8FE4-4524-44B1-B5F8-BCC58D4FDF8E |
|
DOI |
https://doi.org/10.5281/zenodo.3796974 |
|
persistent identifier |
https://treatment.plazi.org/id/0384A837-8267-FFE2-FF37-FD40FE2B7D41 |
|
treatment provided by |
Plazi |
|
scientific name |
Physalia |
| status |
|
Physalia View in CoL post-Haeckel
At last, we can return to sanity! Or, to paraphrase Merrill (1949, referring to Rafinesque, as quoted by Endersby, 2009, p. 170): “in taxonomy and nomenclature we would have been infinitely better off today had Haeckel never written or published anything appertaining to the subject”. It did not take long for Claus (1889a, b), the latter being an English translation of the former, to give a critical review of Haeckel’s 1888 contributions. Claus particularly took issue with Haeckel’s nomenclature for the parts and appendages of siphonophores. However, in actuality that probably had more to do with the translator than Haeckel himself, since the original text was written in German. Claus wrote a great deal in regard to Haeckel’s Medusome Theory, which he debunked very successfully and regarded it as “fiction”! Indeed it attracted few followers. With regard to Haeckel’s classification of the cystonects, Claus (ibid. p. 196-7) stated: “There are numerous novelties in connexion with the division of pre-existing genera into two or more, and, indeed, on the ground of trifling distinctions scarcely applicable as generic characters. As examples may be cited the division of Physalia into Physalia and Caravella and of Alopleota [sic] into Aloploeota [sic] and Arethusa as also the establishment of two subfamilies associated therewith; further the breaking up of Rhizophysa by its different species into the genera Aurophysa , Cannophysa, Linophysa , Nectophysa , Pneumophysa , and Rhizophysa , and the distinction of two subfamilies as Cannophysidae and Linophysidae upon differences which perhaps justify generic separation”. Indeed, Claus (ibid., pp 195-6) had earlier remarked: “That Haeckel makes a very extensive, indeed almost unlimited, use of his skill in making new and suitable names, is certainly intelligible from the fact that he possesses this faculty in a very high degree and has developed it, by many years’ practice, into a speciality, in which at present no other naturalist can hope to equal him”!
Chun (1897) was one of the last 19 th century reviewers of the family Physaliidae, He , like Huxley, drew attention to a couple of the main offenders, Tilesius, Olfers and Lesson, who had done so much to muddle the already confused taxonomy of the family; and, correctly, also added Louis Agassiz (1862) to this list but, being an adherent to Haeckel’s Medusome Theory, he was reticent to criticise that particular author. However, Chun clearly did not consider that Haeckel’s division of the family Physalidae into sub-families, dependent on the presence or absence of a crest, was valid. In fact he believed that there were only two valid Physalia species; one living in the Atlantic and the other in the Indo-Pacific. The main difference between the two species was the other Haeckelian character, in that the Atlantic form had numerous main tentacles, while the other had only one. So Chun, like Huxley (1859), adopted the specific name for the Indo-Pacific species, Physalia utriculus (La Martinière) [but actually, Gmelin, see above]. For the Atlantic species Chun applied what he considered to be the principles of priority and gave it 52 The name P. pelagica was first used by Lamarck (1801, p. 356). Bosc (1802, p. 166) used the misspelt form P. pelasgica . Haeckel (1888a) attributed this name to Osbeck (1757), but that is pre-Linnaean.
53 Haeckel (1888b) used Tilesius (1813) as the authority although, as discussed above, it appears to be Tilesius (1810).
54 Haeckel (1888b) quotes Eschscholtz (1829) as the authority for Physalia utriculus , but Gmelin (1788, p. 3155) was the first to use the specific name for Medusa utriculus (see above).
55 Although Haeckel (1888b) quotes Lamarck as the authority, but in brackets he says “Péron ... pl. xxix, fig.1. Lesueur & Petit (1807) should be the authority.
56 O.F. Müller (1776) is the original authority for this species, under the name Medusa caravella .
the name P. Arethusa , based on the description and name given by Browne (1756). However, that was contrary to the then emerging International Rule of Nomenclature, as Schneider (1898, p. 190) 57 pointed out when he said: “Linnaeus established in the 10 th edition of Systema Naturae, the Atlantic Physaliid as Holothuria physalis. It follows, therefore, that as a species name physalis , but not arethusa or Caravella , is preferable over both. Chun (97, p. 87) violates the principles of priority (which he believed to be following), when he designated our form: Physalia arethusa Browne (see also the rules of nomenclature).” Thus, after 140 years since it was first named by Linnaeus, Schneider finally established the name Physalia physalis by which the species is known today.
Although Schneider (1898, p. 87) 58 also retained the name Physalia utriculus for the “smaller and less luxuriously developed” Indo-Pacific form, he commented (ibid.) 59: “If there are no morphological differences in the construction of the individual appendages between the two forms, it seems to me that it should be understood that the Pacific form is only a variety of the Atlantic one.” As noted above, it was clear that Schneider was not a friend of Chun, judging by their comments about each others’ work, some of which are definitely worth reproducing. For instance, Schneider (1898, p. 51) 60 began his review of the systematics of siphonophores by stating: “For the present communication I am induced by the two Chun works of 1897 [a, b]. To my regret, I have found that Chun, when judging my second communication of 1896, ‘Grundriss der Organisation der Siphonophoren,’ had very little objective preoccupation, and spoke so disparagingly about many points - without thereby substantiating his opposite opinion or my opinion against earlier statements to acknowledge the grounds for refutation - that in the interest of my own work I consider a detailed criticism of Chun’s recent communications to be useless.” However, he was more forthright (ibid., p. 162) 61 when he stated: “I cannot refrain from expressing my satisfaction that Chun now finally recognizes the extra-capsular attachment of the stinging tube in the young nematocyst. Chun still said ‘92 on p. 156 against my statement of ‘91: ‘I regret having to contradict almost all of Schneider’s information.’ Perhaps the time will come when Chun will no longer have to contradict the information I have provided. Incidentally, a statement of the incorrectness of my findings is just as welcome as a confirmation, because I work in the interest of science. Only then must the inaccuracy be ‘explained’ in detail; empty sarcastic phrases are not enough in scientific studies.” Schneider (1898) gave a quite detailed account of the morphology of rhizophysids, particularly with regard to the structure of the pneumatophore and the gonodendra, and made many comments, not all positive, about Chun’s (1897) description, and, with regard to the giant cells of the pneumatophore, he stated (ibid., p. 167) 62: “The interpretation that Chun presents for the giant cells appears completely untenable to me. He regards them as “buffers,” that “prevent vigorous contractions of the stem and the pneumatocodon above it from rupturing the pneumatophore.” If there is any element that would appear to be inappropriate to fulfil such a function it is precisely these giant cells, 57 Original text: “Linné führt in der 10. Auflage des Systema naturae die atlantische Physalie als Holothuria physalis auf.Als Speciesnamen ergiebt sich summit physalis , nicht aber arethusa oder caravella , die beide bevorzugt werden. Chun (97 p. 87) verstösst gegen die Grundsätze der Priorität (denen er doch zu folgen glaubt), wenn er unsre Form: Physalia arethusa Browne benennt (siehe darüber die Nomenclaturregeln).”
58 Original text: “kleiner und weniger üppig entwickelt”
59 Original text: “Falls nicht morphologische Differenzen im Bau der einzelnen Anhänge zischen beiden Formen bekannt werden sollten, scheint mir die pacifische Form nur als Varietät der atlantischen aufgefasst werden zu können.”
60 Original text “Zu der vorliegenden Mittheilung Averde ich durch die beiden Chun’schen Arbeiten von 1897 veranlasst. Ich habe zu meinem Bedauern constatieren müssen, dass Chun bei Beurtheilung meiner zweiten Mittheilung von 1896: »Grundriss der Organisation der Siphonophoren «sehr wenig objectiv vorgieng und sich über manche Puncte derart abfällig aussprach - ohne dabei seine entgegengesetzte Auffassung zu begründen oder meine gegen frühere Angaben vorgebrachten Gründe einer Widerlegung zu würdigen - dass ich im Interesse meiner eigenen Arbeit eine ausführliche Kritik der Chun’schen neueren Mittheilungen für nüthig erachte.”
61 Original text “Ich kann hier nicht unterlassen, meine Genugthuung darüber auszudrücken, dass auch Chun nun endlich die extracapsuläre Anlage des Nesselschlauches in den jungen Nesselzellen anerkennt. Sagte doch Chun noch 92 auf p. 156 gegenüber meinen Angaben von 91: »Ich bedaure, fast sämmtlichen Angaben von Schneider widersprechen zu müssen.« Vielleicht wird auch die Zeit kommen, wo Chun den von mir 96 gemachten Angaben nicht mehr widersprechen zu müssen bedauert. Übrigens ist mir eine Darlegung der Unrichtigkeit meiner Befunde ebenso willkommen, wie eine Bestätigung, denn ich arbeite ja im Interesse der Wissenschaft. Nur muss die Unrichtigkeit dann auch eingehend »dargelegt« werden; leere sarcastische Phrasen genügen nun einmal bei wissenschaftlichen Untersuchungen nicht.
62 Original text: “Ganz unhaltbar erscheint mir die Deutung, welche Chun für die Riesenzellen aufstellt. Er betrachtet sie als »Puffer«, die »bei energischen Contractionen des Stammes und des ihm aufsitzenden Luftschirmes
ein Sprengen des Luftsackes verhüten«. Wenn irgend welche Elemente dazu ungeeignet erscheinen müssten, so sind es gerade die Riesenzellen, die von äusserst zart spongiöser Beschaffenheit und ausserdem nur von einer sehr dünnen Stützlamelle umgeben sind. Mit demselben Recht könnte man Blätterteig als Puffer zwischen Eisenbahnwagen für geeignet halten.”
which are extremely delicate and of a sponge-like texture and, in addition, are surrounded only by a very thin supporting lamella. With the same reasoning one could argue that puff pastry is suitable as a buffer between railway carriages”.
Schneider (1898) was possibly the first person to make a detailed study of the development of the “cormidial” units in Physalia . On his most developed specimens, he found 11 “cormidial” groups, five in the oral zone, and six in the main one. With regard to the gonodendra, he noted (ibid., p. 187) 63: “The findings generally show that the genital groups are being detached in toto. In addition to the tentacle and its associated polyp you will find, if the genital group is no longer detectable, a short rounded stump, to which the genital group was originally attached. Since, as is well known, no medusa bud on Physalia itself reaches maturity, and rather here germ cells are completely lacking, thus the detached genital group seems to be the site of maturation, and in the future, in order to find more advanced stages, one must look for free-swimming groups when a swarm of Physalia occurs.” As Schneider noted male gonophores on the gonodendra he examined, one presumes, therefore, that he considered both the male and female gonodendra to be detached before both the special swimming bells and the gonophores became mature.
Steche (1910) made a much more detailed study of the order of development of the various “cormidial” groups, based largely on the same material that Chun had looked at. He followed Schneider (1898) in designating the Atlantic species, Physalia physalis , but was adamant that it was distinct from P. utriculus , mainly on the more complex nature of the arrangement of the various zooids in the former, and that there were marked differences in the development of the young zooids. He reviewed the other studies on developing larvae by Huxley (1859), Chun (1887) and Schneider (1897). He criticised the description by the last of these authors on several counts. In one instance he considered that Schneider’s conclusion that the ampulla at the base of the tentacle was very similar to the basigaster of a physonect gastrozooids, since in contained innumerable numbers of developing nematocysts, and which Schneider considered to have separated itself from the gastrozooid in order to give greater freedom to the tentacle, as his findings gave no support to that idea although it is not quite clear what these findings were. He also noted that the “cormidial” groups did not arise in a linear order and, thus, (ibid., p. 369) 64 he concluded: “This proves that at an early stage germinative material must be distributed over the entire stem zone, which is then activated one after another”.
Bigelow (1911) followed Steche in recognising the two species, Physalia physalis in the Atlantic, and P. utriculus in the Indo-Pacific. He pointed out that although the Pacific species, with its single main tentacle, resembled the immature stage of the Atlantic species; nevertheless of all the specimens from the Pacific that he, and previous authors, had examined none possessed more than one main tentacle. However, a year earlier Kawamura (1910), in a somewhat obscure paper in Japanese, had already shown that of his specimens from near Izu Oshima, Japan, two had two major tentacles while one had four. Kawamura, thus, considered utriculus to be a variety of P. physalis .
Moser (1925), having studied a large collection of physaliid specimens from the Gauss and other expeditions, came to the same conclusion in that the Atlantic and Indo-Pacific forms were merely varieties of one species, which would thus be called Physalia physalis . Nevertheless, she still recognised the possibility of two varieties that she referred to as the Atlantic arethusa and the Indo-Pacific utriculus .
The final nail in the coffin for the idea that there were two species of Physalia should have come from Okada (1932, 1935) who examined a large number of Physalia specimens from Japan with a pneumatophore ranging in length from 2 to 100 mm. He also made very detailed studies of the order of origin of the various zooids and, helpfully, used the same system of annotations as Steche (1910) had used. Unlike Steche, however, he finally concluded from his studies (ibid p. 19) 65: “It is wrong, as was believed until now, that the Indo-Pacific Ocean Physalia has only one main tentacle, and that is because we had never observed a fully developed Physalia . The specimen, whose pneumatophore reached 95 to 100 mm long, has as many long and major tentacles as the species living in the Atlantic Ocean.”
63 Original text “Die Befunde lehren ganz allgemein, dass die Genitalgruppen in toto abgestossen werden. Neben dem Fang- faden und seinem zugehörigen Polypen findet man - falls die Genitalgruppe nicht mehr nachweisbar ist - einen kurzen abgerundeten Stummel, an dem ursprünglich die Genitalgruppe festsass. Da nun, wie bekannt, keine Medusenknospe an der Physalia selbst zur Reife gelangt, alle vielmehr hier der Keimzellen vollständig entbehren, so scheint die abgelöste Genital- gruppe die Reifungsstätte zu sein, und man wird künftig, um weiter vorgeschrittene Stadien zu finden, bei Auftreten eines Physalia-Schwarmes nach frei schwimmenden Gruppen suchen müssen.”
64 Original text “Dies beweist, dass schon frühzeitig Keimmaterial über die ganze Stammzone verteilt sein muss, das nachein- ander aktiviert wird.”
65 Original text: “C’est à tort que l’on a cru jusqu’à présent que la Physalie de l’Océan Indo-Pacifique n’avait qu’un tentacule principal, et cela parce que l’on n’avait jamais observé la Physalie en parfait développement. Le spécimen dont le pneu- matozoïde atteint 95 à 100 mm. de long, possède de longs et nombreux tentacules principaux, comme l’espèce vivant dans l’Océan atlantique.”
Totton (1960, p. 304), who made very detailed studies of the budding patterns of the “cormidial” groups, was clearly not particularly impressed with the studies on this subject, made by earlier authors when he stated: “The methods employed had to be purely exploratory to begin with, because previous accounts of the arrangement of the groups of buds in Physalia were too superficial and illustrated either not at all ( Schneider, 1896) or so inadequately ( Steche, 1910; Okada, 1932, 1935) that they could not be checked and built upon. In fact it was necessary to start de novo ”. However, he did commend Steche (1907) for possibly being the first person to recognize that the gonophores, usually assumed to be always male, while the special nectophores were assumed to be the female, could also be female; and that all gonophores on a single gonodendron were of the same sex. Was this the first recognition that cystonects were dioecious? It is not clear.
Totton (1960) also was adamant that there was only one Physalia species, namely P. physalis . In fact, all major reviewers since Bigelow (1911) have recognised only the one species. However, there are still a few exceptions to be found in the literature. Collins (2002) used material from an Atlantic (as P. physalis ) and a Pacific (as P. utriculus ) specimen in his analyses of the molecular phylogeny of the Medusozoa. In one instance (ibid. Fig.1 View FIGURE 1 ) he found that that the “Bootstrap indices under maximum parsimony and minimum evolution criteria” for distinguishing the species were both 100; indicating that there was a high degree of likelihood that those two specimens were more closely related to each other than to anything else included in the analysis. However, although the likelihood is high, one cannot use these data to prove that one is dealing with a single species (Dr Casey Dunn, personal communication).
As Dr Dunn has also commented that the more recent papers that have suggested that there is more than one Physalia species have largely been peripheral to the core siphonophore literature, as for more than 70 years siphonophore systematists have agreed that there is just one. For instance, at least two teams of medics have suggested ways to distinguish more than one species of Physalia . Fenner et al. (1993) suggested that the response to the experimental exposure of Physalia tentacles, from different regions of Australia, to vinegar indicated that there must be at least two Physalia species, although they do not appear to provide any statistical evidence for this conclusion. They (ibid., p. 500) continued with the description of: "A newly differentiated specires of Physalia physalis "! This “species” was differentiated from the Atlantic P. physalis by its smaller size, up to 15 cm, and the number of "main" tentacles, up to five, with up to ten developing ones. They then stated (ibid.): "We suggest that the smaller, single-tentacled jellyfish commonly known as the "bluebottle" should be referred to by its original name Physalia utriculus , despite the suggestion in 1960 by Totton that all Physalia species are the same and should all be referred to as Physalia physalis . (This question has remained taxonomically unresolved, and Totton’s suggestion has since been ignored by some workers in this field.)”; this field being medicine!
Before continuing the discussion on Australian Physalia species, let us consider the other team of medics mentioned above. Yanagihara et al. (2002) studied the cnidome of specimens of a Physalia caught off Hawaii, and compared their data with that of previously published information on P. physalis . They noted (ibid. p. 140) that, for the latter species: “Totton & Mackie (1960) … reported large and small holotrichous isorhizas along the tentacle, isorhizas along the stomach lip-region and stenoteles in its gastrozooids and dactylozooids palpons and float.” However, Weill (1934) had identified the isorhizas as atrichous, while others variously had identified them as anisorhizas; and the stenoteles as euryteles. Yanagihara et al. (ibid.) considered that: “Accurate identification of the cnidome is essential in the analysis of venoms and understanding the potential plasticity of predator-prey relationships” but clearly they also thought that it had taxonomic importance, as they were convinced that their specimens belonged to P. utriculus . They made detailed measurements of the discharged threads of the tentacular nematocysts and came to the conclusion that, for their Physalia specimens, they were two sizes of heterotrichous anisorhizas, rather than isorhizas. However, they did not resolve whether the other, non-tentacular, nematocysts where euryteles or stenoteles.
But do the findings of Yanagihara et al. (2002) have any taxonomic significance? It seems, to the present author, that the answer has more to do with semantics than with taxonomy. They found that for both their types of “heterotrichous anisorhizas” the tubule decreased in diameter from base to tip, but that the most rapid decrease in diameter occurred in the distal half of the tubule. From this they concluded that the nematocysts should be considered to be anisorhizas, although they do not state why they came to that conclusion, except to say (ibid. p. 147) that they: “arbitrarily interpreted tapering of more than 30% to be significant”.
Werner (1965, p. 8) described isorhizas as “Faden isodiametrisch”; and anisorhizas as “Faden an der Basis deutlich erweitert”. Similarly, Mariscal (1974, p. 136) defined isorhizas as “thread of the same diameter throughout”; and anisorhizas as “thread slightly dilated toward base”. Östman (2000, p. 44) gave a very similar definition for anisorhizas, but enhanced the definition of isorhizas as: “tubule isodiametric or nearly isodiametric proximal to the mid-point and tapering thereafter”. Thus, using the data of Yanagihara et al. (2002) one finds that, for the smaller type of nematocyst, the diameter of the thread has reduced to just 89% of its proximal value by its mid-length, but to 50% at it distal tip. For the larger for the values are 88 and 65% respectively. Thus, according to Östman’s definition, they should be considered as isorhizas, and the complete absence of any basal swelling on the tubules attests to that conclusion. Thus, on this basis it appears that the conclusions reached by Yanagihara et al. (2002) do not conform with Östman’s definitions.
Pontin (2009), in his Ph.D. thesis, used CO1 and ITS1 DNA molecular sequencing on tissues from specimens of Physalia collected from various locations in New Zealand. He concluded that there appeared to be at least three species of Physalia present, of which only one was likely to be a named species, namely P. utriculus , and neither of the others was P. physalis . However, in the published version of these results, Pontin & Cruickshank (2012) were more cautious in their conclusions, and considered that there was substantial cryptic diversity amongst the specimens of Physalia from New Zealand coastal waters. Nevertheless, they still considered the possibility of there being more than one Physalia species, unless there was an extraordinary level of intra-specific diversity for P. physalis .
Unfortunately, Gershwin et al. (2010) again used morphological characters, such as size and the number of tentacles, to establish that there were four different forms of Physalia in Australian waters. Three of these possessed a single main fishing tentacle, but differed as to whether the crest on the pneumatophore was absent, incomplete or complete. One of these, the commonest of all, the one with a prominent but incomplete crest, they suggested was P. utriculus , with which they synonymised P. megalista ( Lesueur & Petit, 1807) . The other two were thought to have been previously unrecognised species, although it is difficult to believe that a similar one could not be found among the more than fifty Physalia species that have been described to date. The fourth type had multiple tentacles, with or without a crest, but they did not definitively name this as P. physalis . These conclusions do not agree with the conclusion reached by Fenner et al. (2003) who called the last, not the first, P. utriculus . It is difficult to believe these conclusions, in rather the same way as Haeckel’s (1888b) numerous varieties have all been rejected. In addition, their photographs of the various “species” raises the same questions of interpretation with regard to the number of main tentacles as was discussed above with regard to Haeckel’s species.
Bardi & Marques (2007) made detailed morphological and histologival studies of Physalia physalis from the Brazilian waters. The question as to whether the nematocyst batteries tentacles of P. physalis should be considered as tentilla was raised recently by Munro et al. (2018). On the young tentacles these batteries form semi-annular horseshoe-shaped buttons appparently attached directly to the siphosome, but for the mature relaxed tentacles they are arranged along the axis of the siphosome; thereby suggesting that they where ectodermal swellings. However, they noted that Bardi & Marques (2007) had found that the buttons actually contained a cavity, lined with endoderm, that connected with the gastrovascular cavity of the tentacle; thereby suggesting that they were reduced tentilla.
Bardi & Marques (2007) noted that there were regional differences in the morpholoy of their specimens, mainly regarding the number of mature tentacles, which harks back to the results from Australia and New Zealand. They then discussed the possibility that there were more then one Physalia species, but noted that their specimens with more than one main tentacle were longer in length than those with one. Thus, they concluded that the addition of new tentacles was just a stage in the growth of the colony. They also considered the size and types of nematocyst present on the tentacles and noted the results of Yanagihara et al. (2002) that suggested a difference between the Atlantic P. physalis and the Pacific P. utriculus . However, we have already discussed this above and concluded that is just a matter of definition.
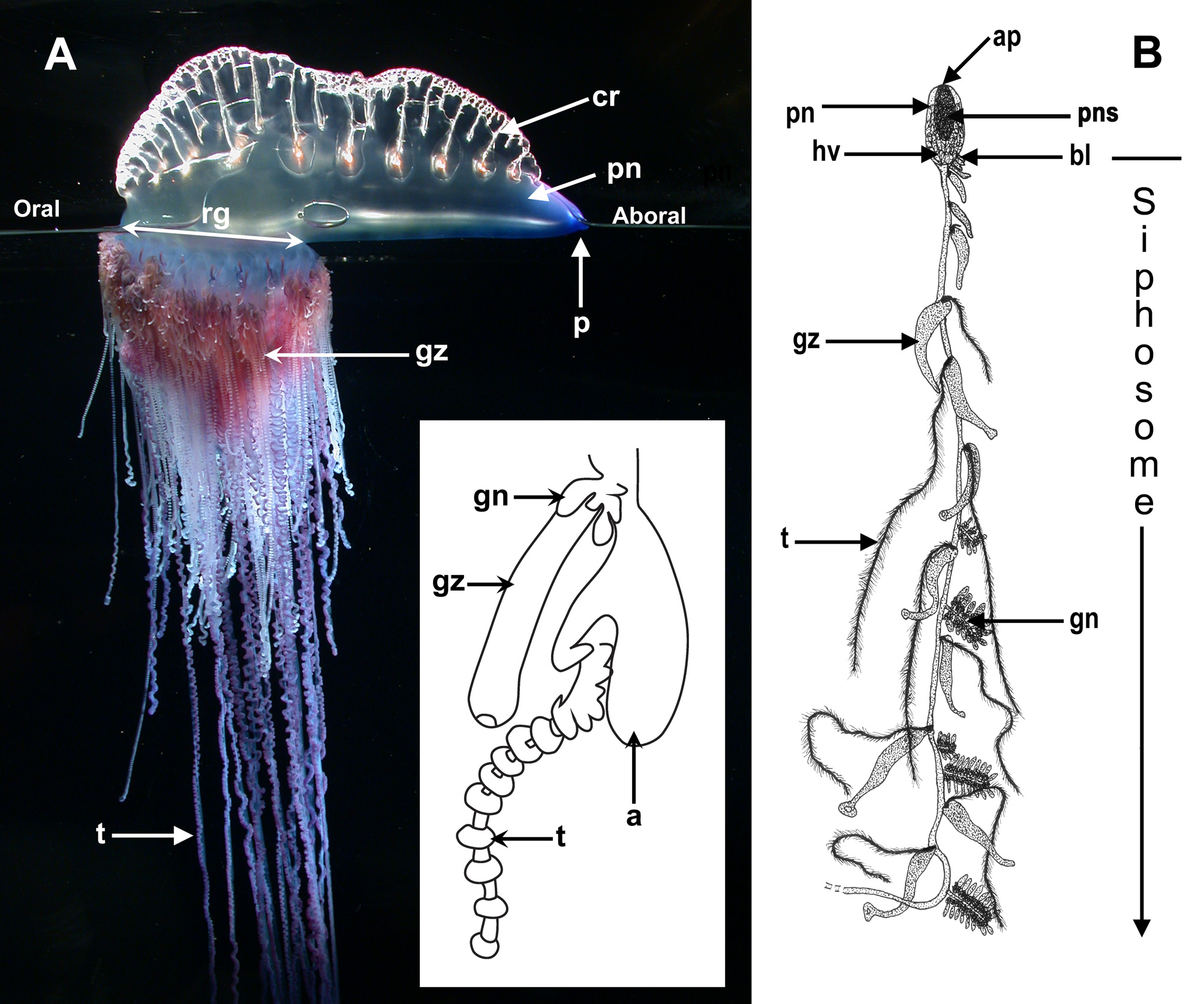
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.