
Reticulammina novaezelandica Tendal, 1972
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4455.3.16 |
|
publication LSID |
lsid:zoobank.org:pub:9FF7EC56-E30D-41E9-A748-D1132F3F07D9 |
|
DOI |
https://doi.org/10.5281/zenodo.5957462 |
|
persistent identifier |
https://treatment.plazi.org/id/038687F5-5354-FFDB-FF08-1599C4D1D184 |
|
treatment provided by |
Plazi |
|
scientific name |
Reticulammina novaezelandica Tendal, 1972 |
| status |
|
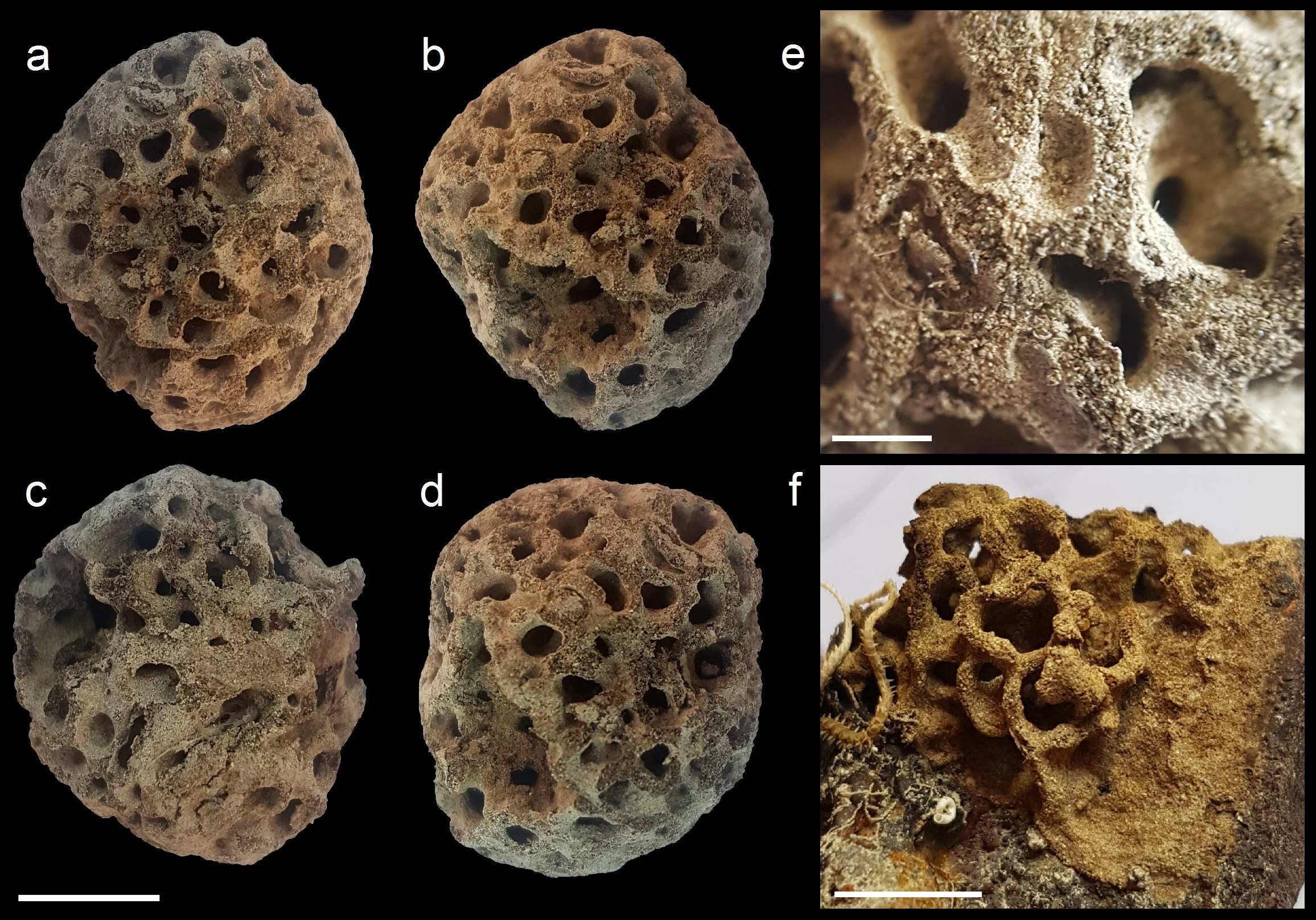
Reticulammina novaezelandica Tendal, 1972 ( Fig. 1A–D View FIGURE 1 )
Reticulammina novaezelandica Tendal, 1972: 29 , pl. 3E–G; Tendal, 1975: 94, fig. 1; Tendal, 1976: 86. Tendal & Lewis, 1978: 202. Dawson, 1992: 83; Tendal, 1996: p. 86. Hayward et al 2007: 265. MacDiarmid et al 2013: 60, fig. 3–23
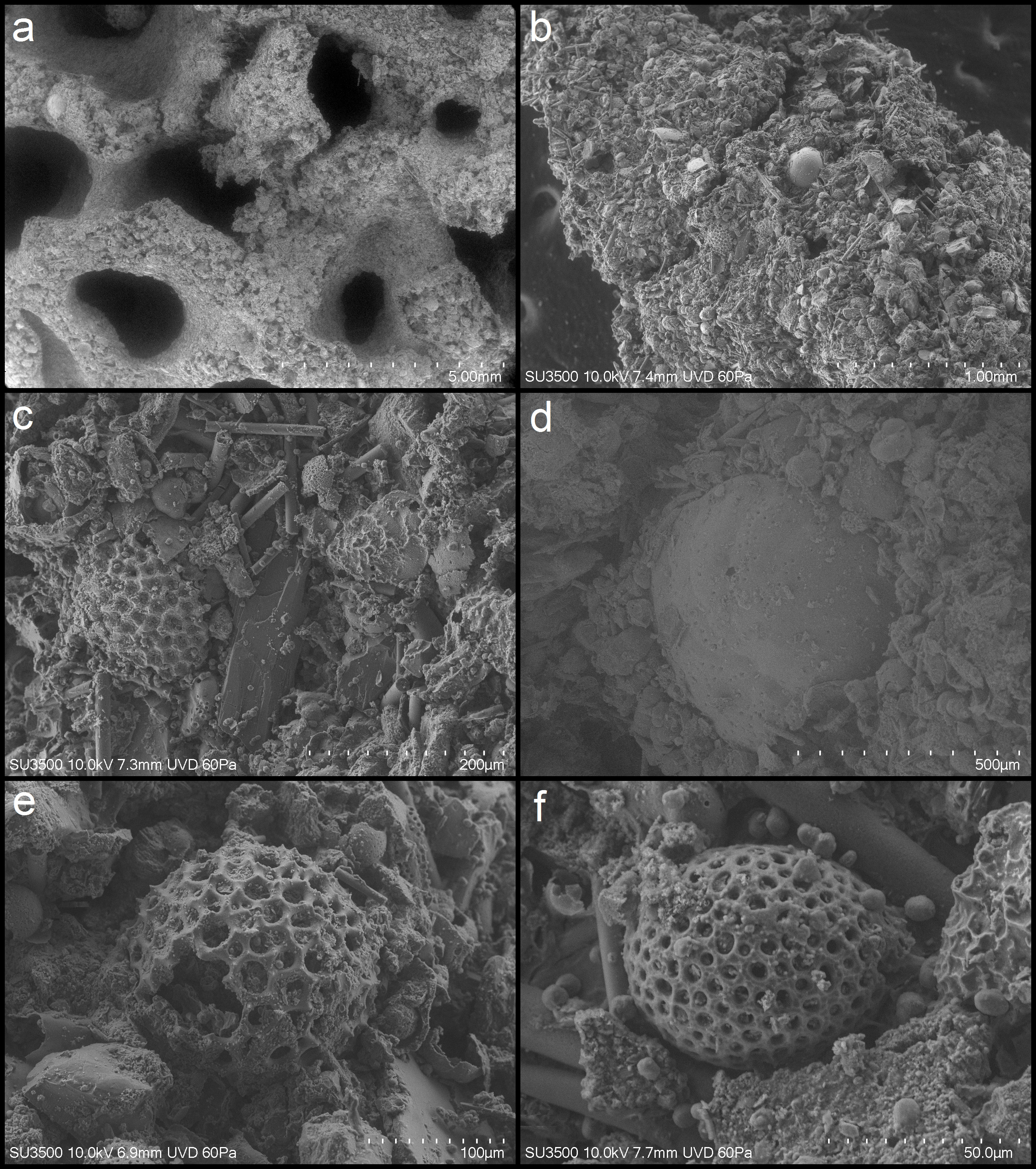
Description. Test rounded, measuring up to about 32 mm in maximum dimension. It is composed of anastomosing lamella-like branches that measure about 0.8 mm to 1.40 mm in width. The open spaces of the test are mostly circular, occasionally oval, and 2 to 4 mm wide. The colour is light greyish-brown. Internal xenophyae are visible on abraided surfaces. They are abundant and consist mainly of foraminiferan and radiolarian tests, with a few sponge spicules and some fine sediment particles. The thin surface layer is composed of fine sediment with a few scattered foraminiferan tests. Granellare and stercomare cannot be seen, but some scattered stercomata, with diameters of 20–80 micrometers, are visible in SEM images ( Fig. 2F View FIGURE 2 ).
Distribution. Reticulammina novaezelandica was described originally from two specimens collected at 960–984 m at the top of Aotea Seamount in the Eastern Tasman Sea west of North Island ( New Zealand), and from a third collected at 743 m on the continental slope off the Bay of Islands, east of North Island ( Tendal, 1972, 1975). The present specimen is the first record for the species outside New Zealand waters.
Remarks. The new specimens closely resemble the original description and illustrations given by Tendal (1972; see also Tendal, 1975) of Reticulammina novaezelandica , leaving little doubt that they represent the same morphospecies. The only obvious differences compared to the holotype ( Tendal 1972, Pl. 3, figs E–G) are that the test is about half the size (32 mm diameter compared to 60 mm), more globular in shape, and the open spaces within it are more rounded. Unfortunately, since DNA data are not available for the Chilean specimen or for any of the original New Zealand material of R. novaezelandica , the identification could not be confirmed genetically, leaving open the possibility that they represent morphologically congruent but genetically distinct cryptic species.
The present record doubles the depth range of this morphospecies (from 743–984 m to 1800–2000 m) and extends its geographical distribution by approximately 128° (more than 10330 km) in an easterly direction across the South Pacific Ocean. Little is known about the biogeographic ranges of xenophyophore morphospecies. Many that occur in the Pacific (for example, most Stannophyllum species) are currently unknown in the Atlantic, and vice versa ( Tendal, 1972, 1996). Morphological evidence suggests that at least some species have limited ranges across the CCZ, with distributions possibly related to gradients in depth and surface productivity ( Gooday et al., 2017c). On the other hand, wide ranges have been reported for a few species. Reticulammina labyrinthica was described from New Zealand waters ( Tendal 1972) and subsequently from the NE Atlantic ( Gooday and Tendal 1988). Syringammina fragilssima was described from the Scottish margin and subsequently from New Zealand ( Tendal 1975). Stannophyllum globigerinum is recorded from the eastern and central Pacific, around Indonesia, off East Africa and the NE Atlantic, while Cerelasma massa is recorded from off East Africa and the NE Atlantic ( Tendal 1972). An important caveat, however, is that these distributions are based entirely on morphological characteristics. Given the morphological plasticity of xenophyophore tests, and the relatively few characters that are available for discriminating between species, it is important for such wide ranges to be confirmed by genetic data.
Levin and Gooday (1992) hypothesized that xenophyophore species, particularly those with reticulate or folded test morphologies apparently adapted to trapping suspended food particles, are most common on sloped or elevated terrain associated with enhanced current regime and particle flux. This concurs with previous observations of suspensionfeeding macroinvertebrates, including antipatharians, cirripedians, octocorals and pennatulaceans, living in the same fishing grounds as Reticulammina novaezelandica ( Araya et al 2016a, 2016b, 2016c). The presence here of this xenophyophore species is consistent with the high megafaunal diversity observed at bathyal depths off the northern Chilean margin. We suspect that these giant foraminifera will prove to be abundant in this region, as they are on continental margins in other parts of the world ( Tendal and Gooday 1981). Xenophyophores probably play an important ecological role in areas where dense populations occur by creating structures that host assemblages of macrofaunal, meiofaunal and microbial organisms ( Levin and Thomas, 1988), as well as promoting the deposition of organic-rich particles that benefit sediment communities ( Levin and Gooday 1992).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Reticulammina novaezelandica Tendal, 1972
| Araya, Juan Francisco & Gooday, Andrew J. 2018 |
Reticulammina novaezelandica
| Tendal, 1972 : 29 |
| Tendal, 1975 : 94 |
| Tendal, 1976 : 86 |
| Tendal & Lewis, 1978 : 202 |
| Dawson, 1992 : 83 |
| Hayward et al 2007 : 265 |
| MacDiarmid et al 2013 : 60 |