
Macrozamia miquelii, (F.Muell.) A.DC.
|
publication ID |
https://doi.org/10.1016/j.phytochem.2021.112715 |
|
DOI |
https://doi.org/10.5281/zenodo.8276707 |
|
persistent identifier |
https://treatment.plazi.org/id/038787AA-6D7E-2353-912B-FA5FFDDE4496 |
|
treatment provided by |
Felipe |
|
scientific name |
Macrozamia miquelii |
| status |
|
2.3. Variation in volatile emissions of M. miquelii View in CoL View at ENA cones
2.3.1. Temporal patterns of cone thermogenesis and volatile emission rates
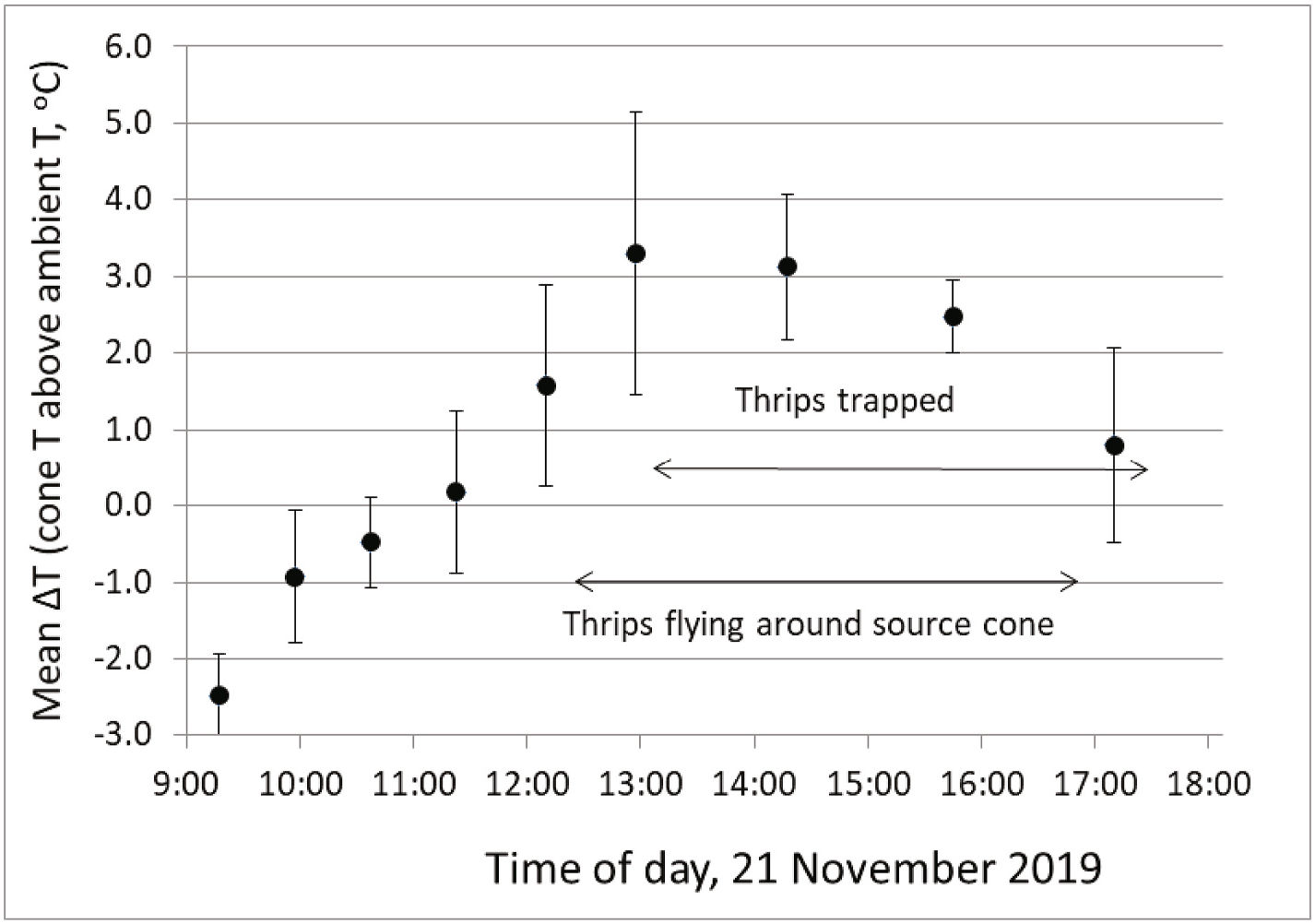
Both field and laboratory studies indicated that both pollen and receptive ovulate cones of M. miquelii exhibit a daily daytime thermogenic event, as in other- Cycadothrips- pollinated Macrozamia ( Terry et al., 2004a) . In the field, where ambient midday temperatures often exceeded 30 ◦ C, pollen cone temperatures reached a maximum during 12:00–13:30 h, ranging from 0.7 to 8.5 ◦ C above the ambient across all pollen cones, with an average of 3.7 ◦ C ( Fig. 7 View Fig ). The figure shows that thermogenic cones passed the ambient temperature around 11:30 h. By 17:00 h, cone temperatures cooled close to the ambient. Experiments on four excised pollen cones under controlled conditions in an environmental chamber also showed a midday thermogenic peak. Cones reached their temperature peak between 11:00 to 13:00 h exceeding the 25 ◦ C ambient by 9–11 ◦ C. On a different day, five receptive ovulate cones reached an average of 5.5 (± 0.77 s.e.) o C above the ambient, measured between11:53 and 12:45 h, while five pre-receptive ovulate cones were not thermogenic with an average 0.16 (± 0.37 s.e.) o C below the ambient temperature measured between 12:08 and 13:10 h.
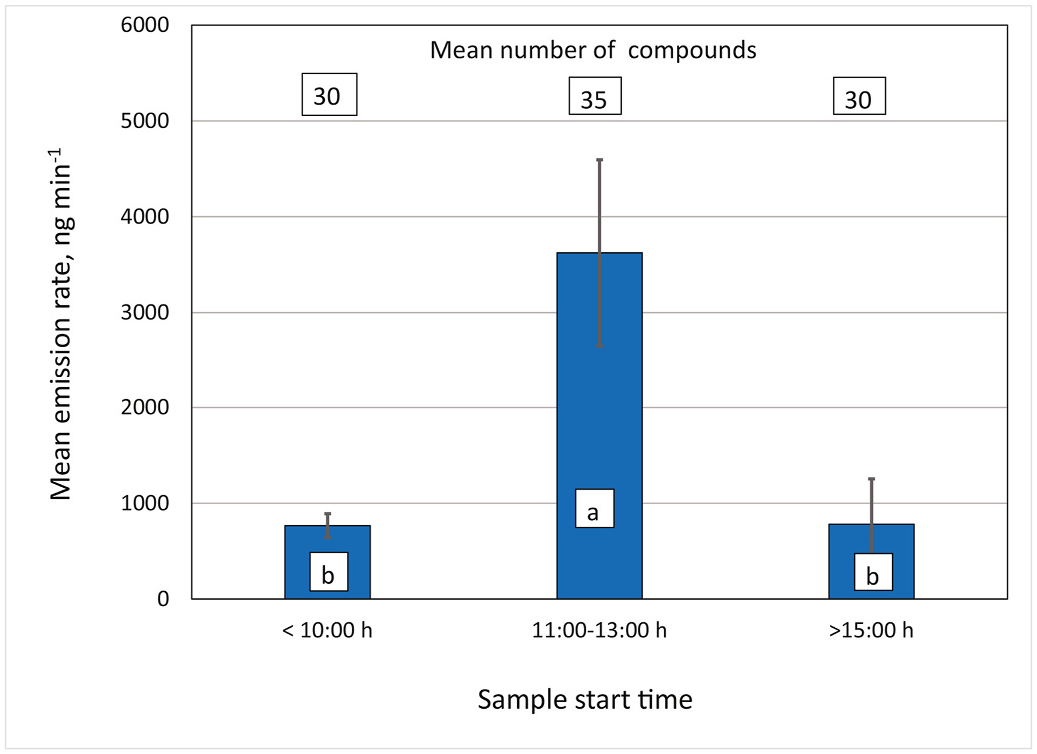
The volatile emission rates of three pollen cones sampled on plants in the field were significantly higher (about five-fold) at midday (during peak cone temperatures) compared with morning or later afternoon rates (ANOVA, F 2,4 = 12.4, P = 0.019) ( Fig. 8 View Fig ). The number of compounds detected from each cone was numerically higher at midday, although the values were not statistically different from one another (Friedman test χ 2 2 = 4.7, P = 0.097). Emission rates of major compounds of each cone also were the highest at midday (Supplementary Fig. S4 View Fig ). Only one receptive ovulate cone was sampled at these same periods in one day, and the midday sample emission rate was five-fold that of the morning and 4.4-fold that of the afternoon sample, with 20, 33, and 23 compounds detected in the early, midday and later afternoon samples, respectively. Another receptive ovulate cone had a midday sample with 2.1 higher emission rate over the morning sample, with 29 versus 24 detected compounds, respectively. In a third ovulate cone, the midday sample had a four-fold higher emission rate over the later afternoon sample, with 33 versus 31 compounds detected, respectively. All subsequent ovulate cone samples were taken during midday to synchronize the sample times near peak thermogenesis.
2.3.2. Variation in monoterpene emission rates among cones
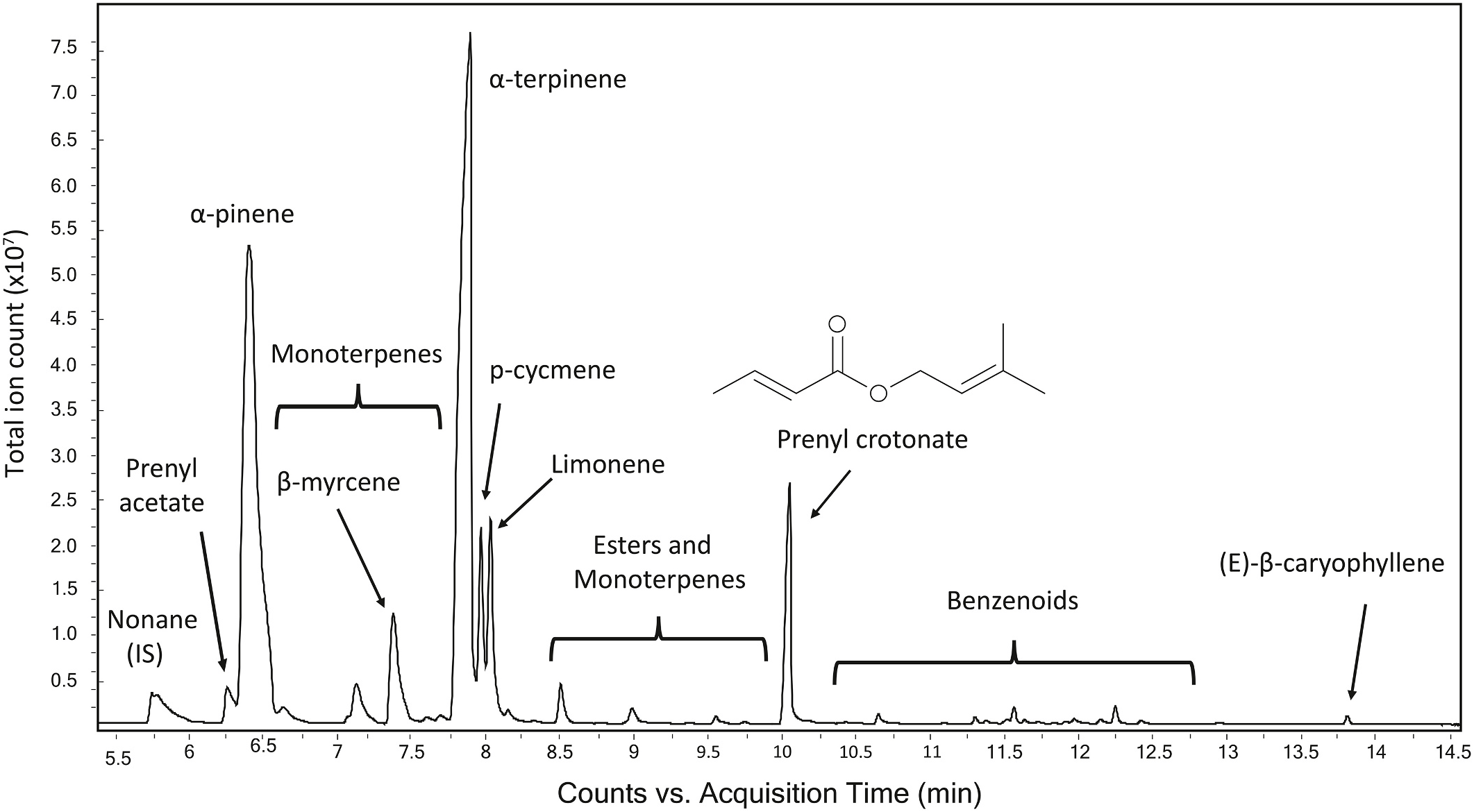
The volatile emission rate across thermogenic events of the same cone (n = 4 pollen cones, each on a different plant, each sampled over six sequential thermogenic days in the field) revealed that the large differences in cone monoterpene emission rates were not the result of differences across cone stage or time of day, but rather to differences across individual cones in their emission rates of several monoterpene compounds. These were mainly α- pinene, α- terpinene and limonene ( Fig. 9a View Fig ), as is evident in the chromatograms of two cone samples that contrast one another in these monoterpene emissions ( Figs. 4 View Fig and 5a). p-Cymene also varied in emission rate among cones but was highly correlated with limonene emission rate (Pearson r = 0.91, P < 0.0001), so it is not included in Fig. 9a View Fig for clarity. All other monoterpenes were minor components except for β- myrcene. Of the four cones tracked over six days, two were high emitters of these monoterpenes (e.g., maximum for α- terpinene, 29,979 and 9733 ng min 1, Cones 1 and 2, respectively), and two barely emitted them (maximum for α- terpinene, 94 and 10 ng min 1, Cones 3 and 4, respectively). These three compounds comprised > 70% of the total volatile emissions from Cones 1 and 2 but ≤4% from Cones 3 and 4. Over the course of all the sample days, these percentages remained close to the same on each cone.
By contrast, emission rates of two other components, MBO and prenyl crotonate, were only about five-fold different across the four cones ( Fig. 9b View Fig ) during the same sample time period, and Cone 3 had the highest mean emission rate of both compounds. These two compounds comprised over 70% of the total emissions from Cones 3 and 4, but ≤7% from Cones 1 and 2. In addition, prenyl crotonate and MBO are present in all four cones at high levels ( > 800 ng min 1 at midday), and emission rates of both compounds increase between morning and midday samples.
Emission rates of β- myrcene, another monoterpene, varied less than ten-fold across the cones (data not shown). Together, these data demonstrate that plant-to-plant differences in emission rates of α- terpinene, α- pinene, limonene and p-cymene monoterpenes by cones is not simply a function of some cones being high emitters of all compounds.
Because two cones emit high levels and two cones very low levels of these four monoterpenes, the pattern could be interpreted as those monoterpenes being produced by the same terpene synthase (TPS). When we examined the emissions of all the pollen and receptive ovulate cones, six of 10 pollen and three of 14 receptive ovulate cones were high emitters of monoterpenes. However, three of the cones that were high emitters of α- pinene ( > 15,000 ng min 1 per sample) emitted very low α- terpinene levels (two cones emitted < 115 ng min 1 and a third emitted ~ 2800 ng min 1) (Supplementary Fig. S5a), demonstrating these two compounds’ production are not always coupled. A similar pattern was found between limonene and α- pinene but there was a tight correlation between limonene and α- terpinene (Supplementary Fig. S5b, c).
2.3.3. Comparisons of volatiles across receptive and non-receptive ovulate cones
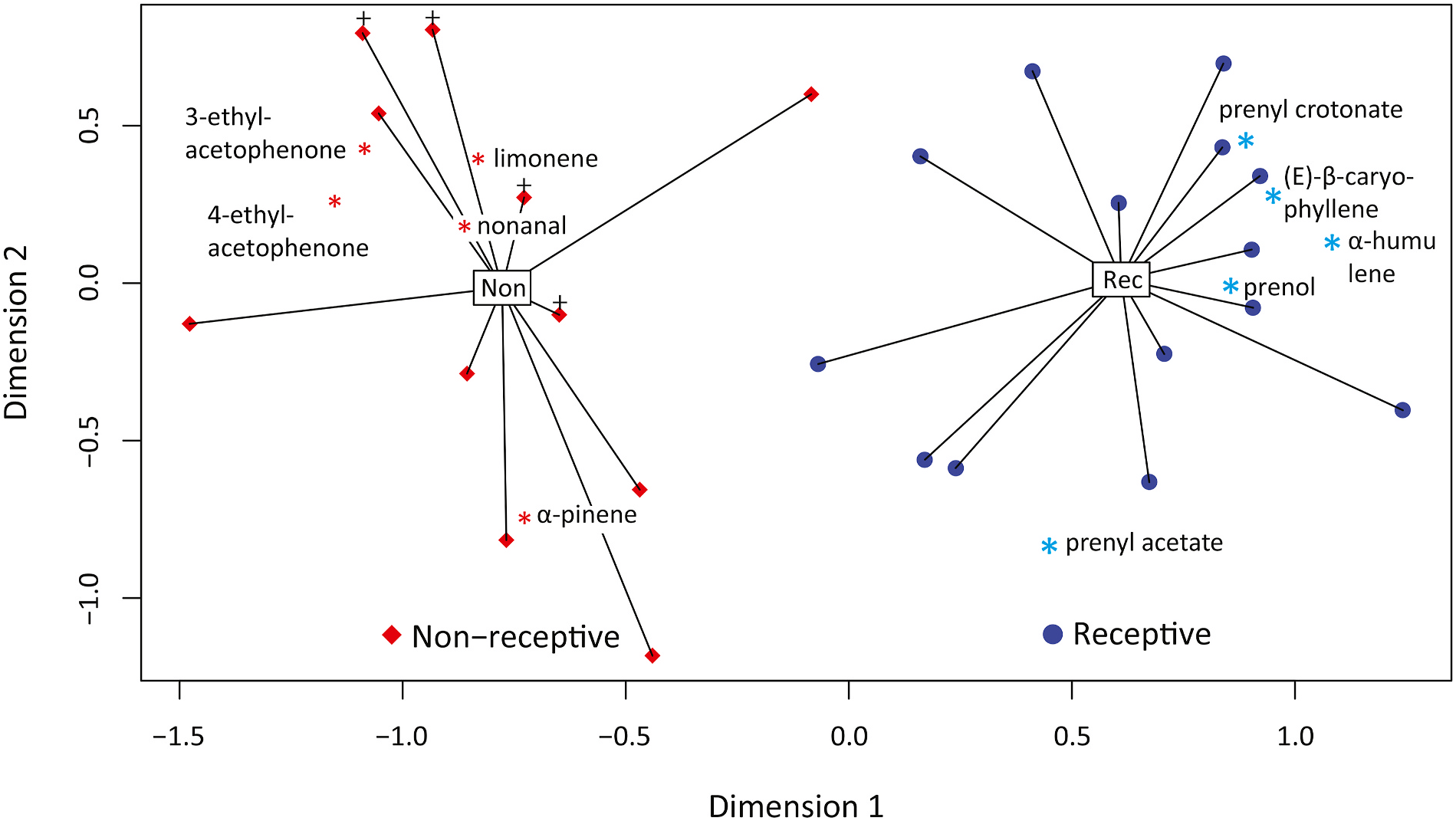
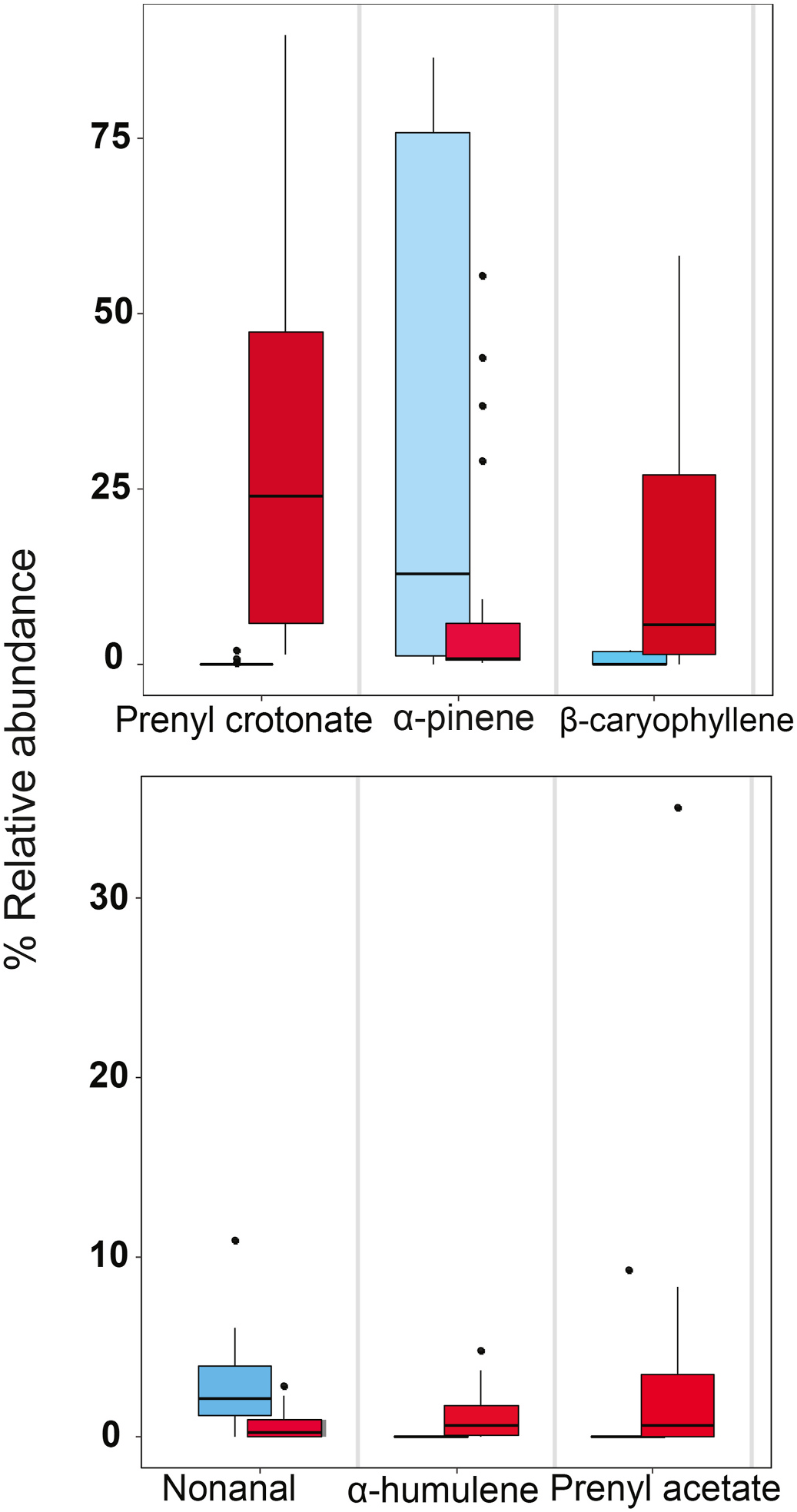
A significant difference was evident across receptive and non-receptive cones in the percentage that each compound contributed to the total volatile emissions. The compounds that contributed differentially across receptive and non-receptive ovulate cones were prenyl crotonate, α- pinene, prenyl acetate, β- caryophyllene, α- humulene, nonanal and limonene ( Figs. 10 View Fig and 11 View Fig ), as well as the two ethylacetophenone compounds. Also, MBO was different across receptive and non-receptive cones. MBO was detected in all receptive ovulate cones at a mean emission rate of 250 ng min 1, but was detected in only one non-receptive cone at trace level ( Table 1 View Table 1 ). It could not be included in the ordination statistical test associated with Fig. 10 View Fig because all samples used in the test were not run on the polar column.
2.4. Attraction of pollinators
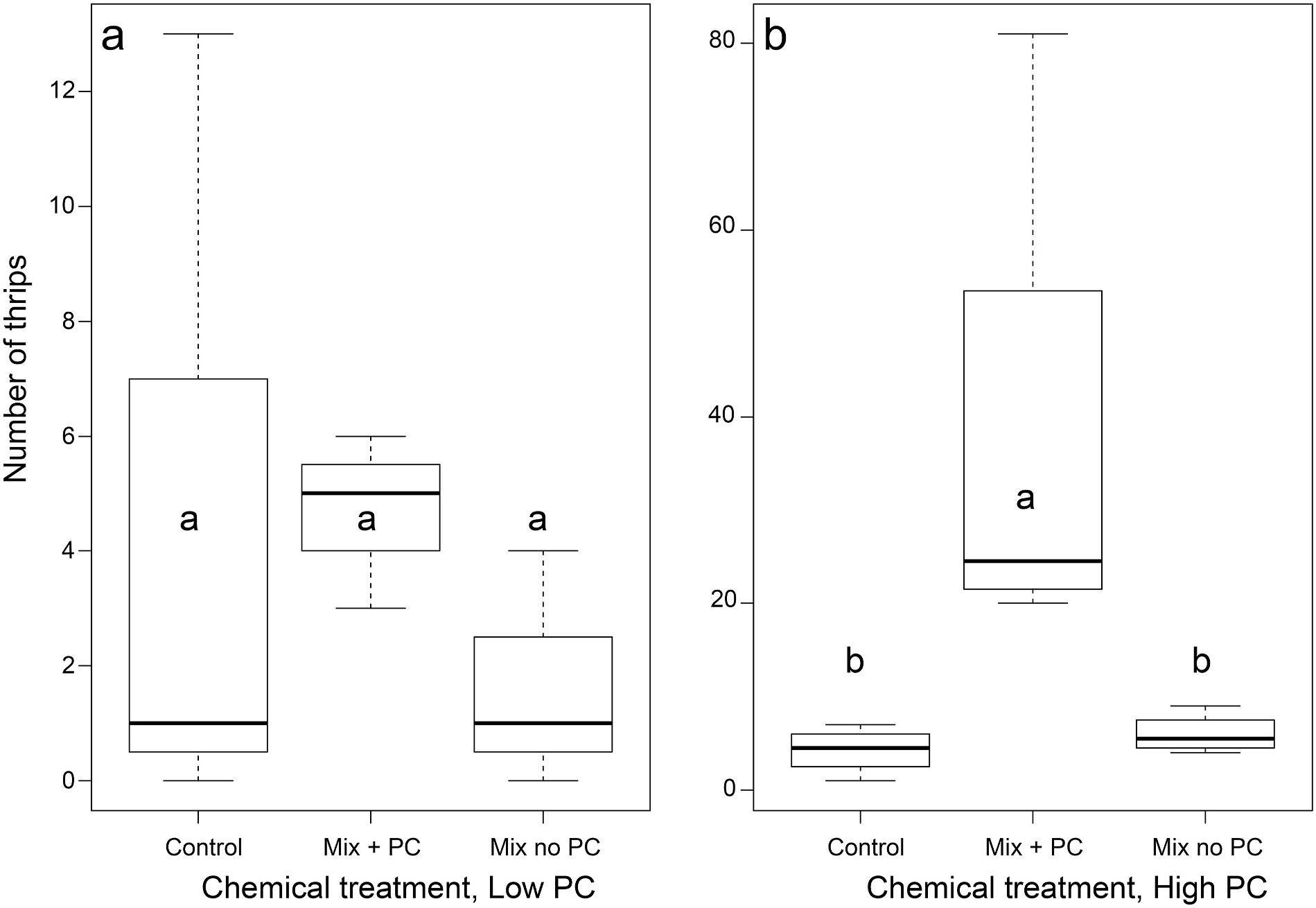
Field bioassays in 2018 indicated that thrips are not attracted to the two compounds tested, prenyl acetate and α- pinene, each one tested at 10 μL per septum. Only two thrips were found on prenyl acetate traps, none on α- pinene traps, and one on the control traps (totals across four replicates and results from tests conducted on two days were combined). In 2019 assays, with a small volume of prenyl crotonate added to the standard bouquet (8 μL per septum), there was no significant difference among treatments in the number of trapped thrips at the end of the experiment ( Fig. 12a, F View Fig 2 View Fig , 6 View Fig = 2.6, P = 0.108)). When more prenyl crotonate was added to the standard mixture in a second experiment (at 40 μL per septum), significantly more thrips were captured on the prenyl crotonate treatment than on the non-prenyl crotonate treatment or the control (treatment effect, F 2,6 = 17.063, P = 0.0013, Tukey’ s HSD, α < 0.05; Fig. 12b View Fig ). Many thrips were observed landing on the source pollen cones in the afternoon, so our treatments were competing with the host cones as well as other cones in the vicinity. The prenyl crotonate emission rate in the latter tests ( 463 ng min 1) was slightly higher than its mean emission rate in the midday receptive ovulate cone samples ( 386 ng min 1, Table 1 View Table 1 ) based on dynamic head space sampling of the bottle treatment apparatus.
In a Y-tube olfactometer experiment, both male and female thrips were attracted significantly to prenyl crotonate at the lower rate [binomial test, probability of going to prenyl crotonate vs control is equal to 0.5: 0.65 vs 0.35 (out of 143 thrips), P < 0.002; 0.67 vs 0.33 (out of 157 thrips), P < 0.002, n = 14 runs for both females and males, respectively with a 95% confidence interval of 0.66–0.998], but not to the higher rate [0.62 vs 0.38 (out of 103 thrips), P = 0.064; 0.55 vs 0.45 (out of 100 thrips) P = 0.39, n = 9 runs for both females and males, respectively), nor with males and female trials combined, P = 0.14, 95% confidence interval, 0.44 to 0.897]. There was no significant difference in response by sex (‘prop test’, χ 2 = 0.4, df = 1, P = 0.835 for 1 μL test; χ 2 = 0.0.0023, df = 1, P = 0.96 for the 5 μL test).
3. Discussion
In the following, we incorporate our major findings into the broader context of other research on cycad and floral volatiles. We deal with the timing of thermogenic events and its association with thrips behavior and volatile emissions, how M. miquelii cone volatiles differ from other cycad (even closely related ones) and plant volatiles, the attraction of C. chadwicki 1 to prenyl crotonate, and the implications for interpreting the evolution and divergence of Cycadothrips chadwicki cryptic species and their host cycads.
3.1. Thermogenesis and thrips pollinator movement
Macrozamia miquelii cones display a daily midday thermogenic event
( Fig. 7 View Fig ), with a concomitant increase in cone volatile emission rates of major compounds ( Figs. 8 View Fig and 9 View Fig ). These results are similar to those of other Cycadothrips -pollinated Macrozamia species in having a daytime thermogenic peak compared with Macrozamia species pollinated by Tranes weevils, which have an evening thermogenic peak ( Terry et al., 2004 a,b). Thus, thermogenic timing is not a feature that differs across the cycad species that host different cryptic species of C. chadwicki . Thrips begin to leave cone cones when thermogenesis begins to peak and continue their movement through the afternoon ( Fig. 7 View Fig ). This pattern of emigration contrasts with that of the thrips pollinators of M. lucida and southern M. macleayi , which leave cones en masse over a 15–30 min period ( Terry et al., 2004b) near the peak of thermogenesis.
This means that we cannot say that this M. miquelii pollination system operates in a ‘push-pull’ manner as in M. lucida and M. macleayi from southeastern Qld ( Terry et al., 2004b, 2007b, 2014). In these southern Qld species, β- myrcene (together with thermogenic heating and associated high humidity) expels thrips from thermogenic cones, and emission rates of β- myrcene are more than 20 times higher than the rates of β- myrcene emission from M. miquelii cones ( Terry et al., 2007a, 2014). Further, M. miquelii thrips were not attracted to β- myrcene at levels emitted by non-receptive ovulate cones (maximum of 73 ng min 1, Table 1 View Table 1 ) nor were they attracted to the standard mix treatments that included β- myrcene ( 368 ng min 1 assessed from head space sampling of bottle trap apparatus). More significantly, in different bioassays, these thrips were not attracted to traps in the field that included M. lucida or M. macleayi cones emitting very high levels of β- myrcene (Terry et al. unpublished). Whether any of the compounds emitted by M. miquelii or the heating of the cones alone is involved with the ‘push’ aspect of pollination is unclear. Only a few cycad studies include sufficiently detailed behavioral observations to determine confidently whether push-pull is part of the pollination mechanism, but it clearly is in Encephalartos ghellinckii and Zamia furfuracea ( Salzman et al., 2020; Suinyuy and Johnson, 2020), and certainly not so for E. villosus ( Suinyuy et al., 2013a) .
3.2. Macrozamia miquelii cone volatiles compared with those of other cycads
Only about 9% of the ~350 recognized species of cycads have had their cone volatile chemistry analyzed, so the published diversity of cycad chemistry is surely not representative. Nonetheless, several common elements in cone odors, as well as unusual findings, have been revealed in this study. Since so many more cycads await investigation of their cone volatiles, we may yet see other species with similar compounds.
1. Both pollen and receptive ovulate cones of M. miquelii emit a similar array of compounds to one another, the dominant compounds are the same across the sexes, and pollen cone emission rates are generally higher than those in ovulate cones ( Table 1 View Table 1 ), as in other Macrozamia and even some species in other cycad genera ( Azuma and Kono, 2006; Suinyuy et al., 2013a; Suinyuy and Johnson, 2018; Terry et al., 2004b; Wallenius et al., 2012). Some receptive ovulate cones of M. miquelii did, however, have a higher emission rate than some pollen cones ( Table 1 View Table 1 ).
2. Almost all cycads, and all Macrozamia species including M. miquelii , produce monoterpenes, with the known exceptions being Z. pumila (now Z. integrifolia L. F.) ( Pellmyr et al., 1991) and Cycas revoluta ( Azuma and Kono, 2006) . All Macrozamia species investigated produce some β- myrcene, but with considerable interspecies variation in the relative levels ( Terry et al., 2004a, 2004b, 2008). M. miquelii also emits sesquiterpenes, as does M. platyrhachis among the Macrozamia species studied, and (E)-β- caryophyllene and α- humulene are also common to both species ( Terry et al., 2008) suggesting they are produced by the same TPS enzyme, as has been reported in some angiosperms ( Booth et al., 2017; K¨ollner et al., 2008.). influences including selection by pollinators, natural enemies, genetic drift, or physiological status, and such variation is not uncommon even within populations (see Delle-Vedove et al., 2017). Examples of intra-population variation have been reported in some orchid floral volatiles, with presence/absence of some compounds across different plants, perhaps because the odorants involved do not have a pollination, defensive or protective function, and are therefore not exposed to selection ( Dormont et al., 2019). Pleiotropic or epigenetic changes could, under such circumstances, affect the biosynthetic enzymes or regulatory genes ( Gang, 2005). This implies that the four monoterpenes involved are not functional in M. miquelii pollination, and results of field bioassays revealed that none these compounds attracted Cycadothrips , when tested alone (α- pinene) or as part of the standard mix.
4. Five of the M. miquelii cone volatile components have not been reported previously from any cycad. Two of them, prenyl alcohol and MBO (3-methylbut-2-en-1-ol and 2-methylbut-3-en-2-ol, respectively), are hemiterpenes. The three others are prenyl esters. Of these, prenyl acetate is found in most pollen and receptive ovulate cones at moderate levels, and prenyl butyrate is a minor component and relatively uncommon ( Table 1 View Table 1 ).
Prenyl crotonate (3-methylbut-2-enyl (E)-but-2-enoate), by contrast, is a major component representing 70–84% of total emissions in many pollen cone volatile samples and up to 67% in some receptive ovulate cone volatile samples. It is one of the few compounds that was detected in all pollen and receptive ovulate cones ( Table 1 View Table 1 ) at moderate to high emission levels in the midday samples (e.g., Fig. 9b View Fig ), whereas this was not the case with the monoterpenes ( Fig. 9a View Fig ). That it was detected at trace levels in post-receptive cones, and then only three of the four, and not detected at all in the seven pre-receptive cone samples ( Table 1 View Table 1 ), implies functionality. Indeed, prenyl crotonate has been confirmed as an attractant for the specific pollinator of M. miquelii , C. chadwicki 1 ( Fig. 12 View Fig ). This information reinforces the interpretation that this volatile has been selected for its chemical mediation role with respect to the specific pollinator of this plant. These results not only substantiate the M. miquelii thrips pollinator being a different species to those associated with M. lucida and southern Qld M. macleayi , C. chadwicki 4, but also suggest that the host plant and pollinator specificity in these Macrozamia species is associated with major shifts in the compounds that mediate the interactions. Investigating these factors in the other species in relevant clades is crucial to determining the basis of their divergence, as elaborated in section 4.
Sesquiterpenes, mainly (E)-β- caryophyllene, have been detected in some species of Encephalartos , Zamia and Stangeria ( Pellmyr et al., 1991; Procheş and Johnson, 2009; Suinyuy et al., 2010, 2012, 2013b). The role of the sesquiterpene compounds remains unclear.
3. Clear plant-to-plant differences in the production of four monoterpenes, α- terpinene, α- pinene, p-cymene, and limonene, were evident, with some pollen cones emitting them at very high rates, and others only at low levels, or even just traces, regardless of cone stage ( Fig. 9 View Fig ). Other major components did not show such extreme variation. Such large intra-population differences in production of compounds have not been reported in other cycads. Intraspecific variation in volatile emissions, both quantitative and qualitative, in plants across populations is common and may be the result of various
3.3. The compounds newly reported from cycads
Prenyl alcohol and MBO, along with isoprene (another, and well known hemiterpene), are derived from dimethylallyl pyrophosphate (DMAPP) through the mevalonate (MVA) or the methylerythritol 4- phosphate (MEP) pathways ( Zeidler and Lichtenthaler, 2001; Zhao et al., 2013; Zheng et al., 2013). Prenyl alcohol is found in a wide range of plants (Good Scents Company, 2020b). MBO has been found in insect gut fungal microbes and in insect sex and alarm pheromones ( Wheeler et al., 1983; Zhang et al., 2012; Zhao et al., 2015). Also, it is produced by aspen, spruce, and pine trees ( Gray et al., 2003, 2011; Zhao et al., 2015), where it has been proposed as having a similar protective function as isoprene. These compounds were included in the mixture of standards in our bioassay experiments and did not attract the thrips pollinator.
Prenyl butyrate is a male aggregation pheromone of the heteropteran Thaumastocoris peregrinus , a Eucalyptus feeder Calvo et al. (2018); Gonzalez´et al. (2012); Groba et al. (2019); Martins et al. (2012). It may be unusual in plants; it was not reported in the extensive review of floral scent compounds by Knudsen et al. (2006). It has, however, been reported since in the volatiles of the Chinese dwarf cherry ( Ye et al., 2017). Prenyl acetate, by contrast, has been reported from many natural sources including plants ( Pherobase, 2021a; Ye et al., 2017). Prenyl acetate did not attract thrips in our field bioassay. The functional significance of these two prenyl esters in M. miquelii remains unknown.
Prenyl crotonate, the other confirmed prenyl ester, has not been reported from any natural source in all of our literature searches ( Knudsen et al., 2006; SciFinder; Web of Science). This compound was reported in a laboratory study of a novel method of ester synthesis ( Yadav et al., 1998), but no spectroscopic proof of structure has been published. A CAS number has been assigned to prenyl crotonate (SciFinder). Thus, our study is the first to report this ester’ s occurrence in nature, to characterize its structure spectroscopically, and to demonstrate a biological function.
Other esters of crotonic acid, the ethyl ester in particular, have been reported as possible attractants of Diptera (e.g., Jayanthi et al., 2012; George et al., 1986), and this ester also features in the volatile signature of more than 150 plant species ( Pherobase, 2021b). In contrast, the prenyl ester of crotonic acid is unique and is one of very few examples of volatile prenyl esters found in nature. Two others are prenyl butyrate and the quite common prenyl acetate, both present in M. miquelii . In addition, we have established the presence of another compound (currently unidentified) very closely related to prenyl crotonate ( Fig. 5b, compound marked ‘*’, and Table 2 View Table 2 ) and strong indications that trace quantities represented by some of the peaks present in Fig. 5b are also related.
4. Conclusion
Our confirmation of a new chemical compound, prenyl crotonate, emitted by M. miquelii cycad cones to attract their thrips pollinator, raises questions relevant to understanding the evolutionary diversification of these thrips obligate pollinators and their plant hosts. How do the other cryptic species of the C. chadwicki complex respond to the cone odors of M. miquelii which contain many of the monoterpenes common to most other Macrozamia cycads? If the cryptic species C. chadwicki 1, alone, is attracted to M. miquelii cone volatiles, that is to prenyl crotonate, this pollination system would represent an example of a single compound species-specific cue involved in a specialist pollinator’ s attraction ( Chen et al., 2009; Raguso, 2008). This cone odorant could, therefore, be involved in the divergence that occurred within the lineages of these two species. If so, is prenyl crotonate a novel compound recently added to the plant’ s genetic odor producing repertoire? Or, is this chemical a relictual volatile cone trait that has been lost in other cycads? If this is so, and prenyl crotonate had originally a non-pollination function, then it may have been subsequently co-opted for pollination ( Armbruster, 1997). Knowledge of the molecular and genetic basis for production of the prenyl esters in these cycads, and an appreciation of any other role they may have in these plants, would be required to help answer these questions.
An understanding of whether there is congruence in the phylogenetic relationships of the pollinator and their Macrozamia hosts is also needed to answer broader questions concerning the diversification of these two taxa. In other obligate pollination mutualisms (fig-fig wasps, yuccayucca moths, and leafflower), coevolution and co-speciation have been rejected as explanations in favor of changes after geographic isolation, or allopatric speciation, and usually underpinned by host shifts ( Hembry and Althoff, 2016). This has also been demonstrated for the five C. chadwicki cryptic species, whose estimated divergence times across the different species range from 1.1 to 7.3 Mya ( Brookes et al., 2015). Furthermore, if each of the cryptic species in the C. chadwicki species complex is attracted to only its own regional host plants in response to species-specific signals, this would provide further evidence of the species status of each and explain the reproductive independence of their cycad hosts, as seen in some orchids ( Dormont et al., 2019; Joffard et al., 2016; Vereecken et al., 2011). Similar odor-related diversification has been noted among Encephalartos cycad cones and their specialist pollinators ( Suinyuy et al., 2012; Suinyuy and Johnson, 2018), suggesting that cycads on different continents have undergone diversification in similar ways to one another.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |