
Rostanga byga
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3745.2.2 |
|
publication LSID |
lsid:zoobank.org:pub:D87FBB64-5DE2-4D19-9338-6E9BE212FAEF |
|
DOI |
https://doi.org/10.5281/zenodo.6146304 |
|
persistent identifier |
https://treatment.plazi.org/id/0387C073-FFB9-630B-FF22-0B05B0135D2E |
|
treatment provided by |
Plazi |
|
scientific name |
Rostanga byga |
| status |
|
Rostanga byga View in CoL Er. Marcus, 1958a
( Figs. 1 View FIGURE 1 G; 15–16)
Rostanga byga Er. Marcus, 1958a: 22, figs. 34–36; Ev. Marcus (1971: 943); Rudman & Avern (1989: 329); Muniaín & Valdés (2000: 1–10, figs. 3B; 4A–C; 5; 6A–E, tbl.1); Valdés et al. (2006: 182); Schrödl & Grau (2006: 8); Rios (2009: 427); Padula et al. (2012: 3, fig. 6H).
Type material. Holotype AMNH 3507, May 1956, 11.0 mm preserved length.
Type locality. Ilha de São Sebastião, São Paulo state, Brazil.
Material examined. Brazil: Rio de Janeiro state: Cabo Frio: Ilha comprida: MNRJ 13299, 29 /iii/2008, V. Padula coll. [1; one dissected]; Arraial do Cabo: Praia do Forno: MNRJ 11765 14/vii/2007, J. Alvim coll. [1; one dissected]; MNRJ 12371, 23 /iv/2007, J. Alvim coll. [1]; MNRJ12805, 16 /v/2008, V. Padula coll. [3; one dissected].
Geographical distribution. Bermuda, Bahamas, Mexico, St. Lucia (Valdés et al. 2006); Brazil: Maranhão state (Ev. Marcus, 1971); Alagoas state: Saco da Pedra (Padula et al., 2012); (Rio de Janeiro state: Cabo Frio, Arraial do Cabo (present study); São Paulo state: Ilhabela (Er. Marcus, 1958a); Argentina (Muniaín & Valdés, 2000).
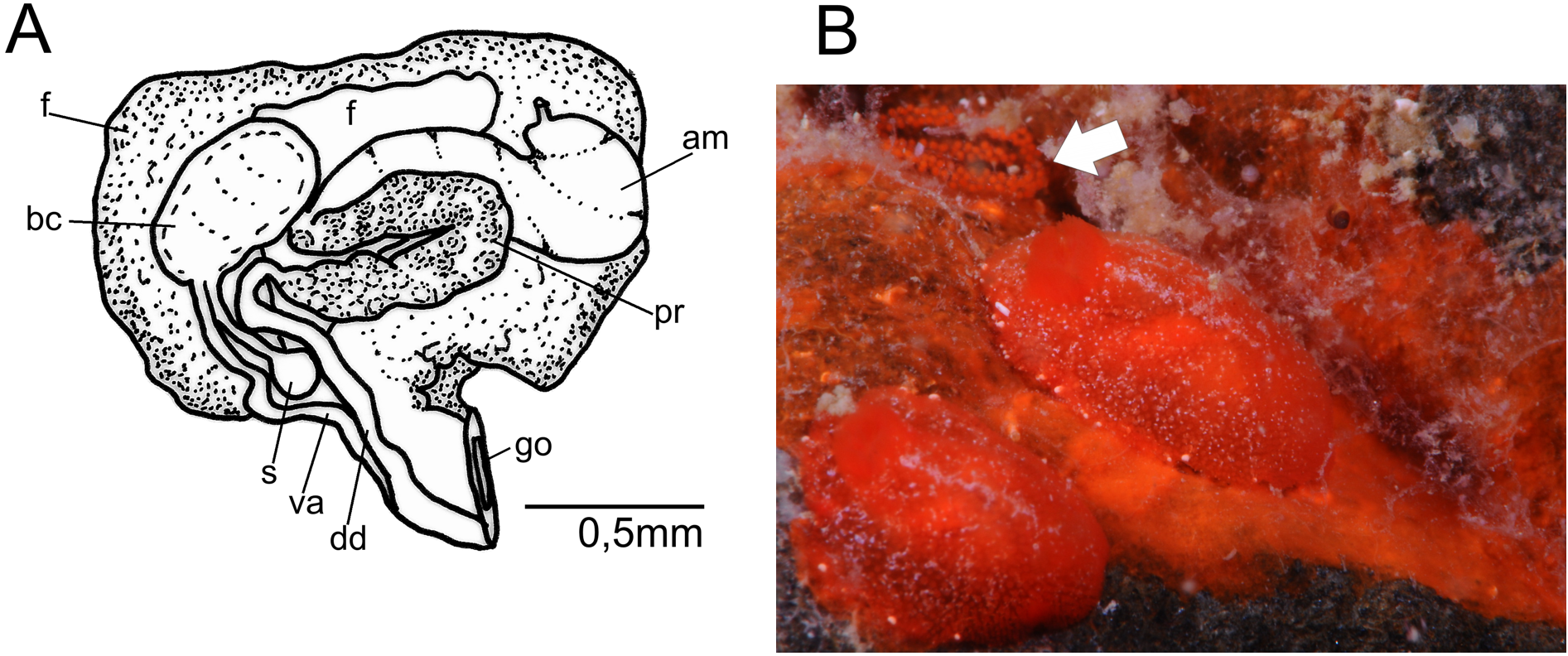
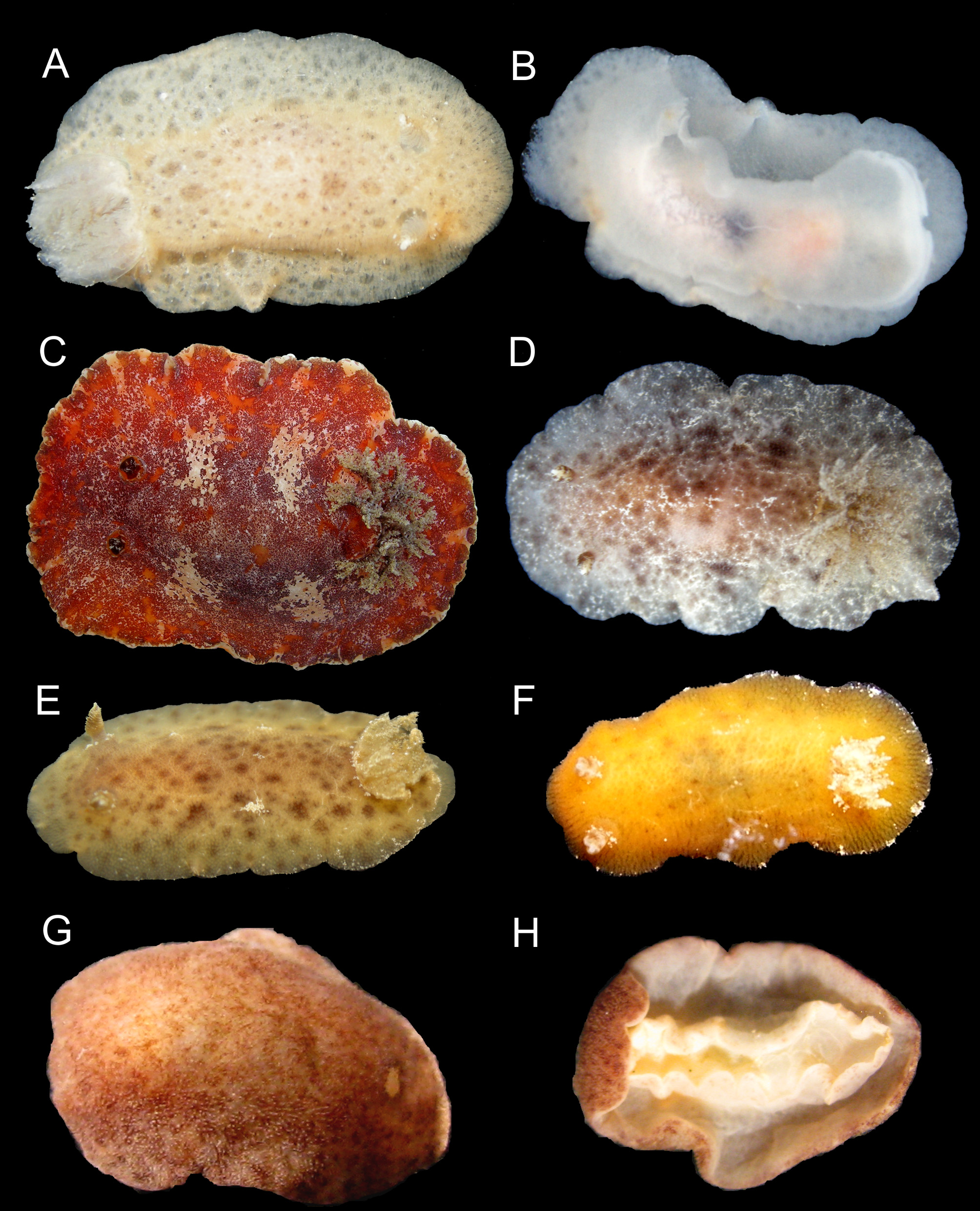
Description. External morphology ( Figures 1 View FIGURE 1 G; 15E–F): body oval, dorsally convex, up to 7.0 mm long alive. Mantle densely covered by caryophyllia approximately of same size and equidistants; with 3–7 spicules that protrude around tubercle; tubercle apex with a ciliary tuft elongated and oval. Rhinophoral and branchial sheaths densely covered by caryophyllidia. Low and wide rhinophores with nine vertical perfoliations, like a rosette with a terminal lobe toward inside of body. Gill with five-six simple branchial leaves, which form a closed circle around high anal cone. Foot narrower than mantle; anteriorly, bilabiated and notched on upper “lip”. Rectangular oral tentacles. Color of living specimens predominantly reddish (brick red), including rhinophores, gill and foot; some specimens present a white longitudinal line on middle of dorsum; in almost all specimens were observed white spots (glands) on edge of mantle approximately equidistant.
Labial cuticle and radula ( Figures 15 View FIGURE 15 A–D): Labial cuticle formed by a smooth cuticle with elements concentrated in two areas on median portion of plates; elements are arranged in about eight rows with about 23 well-marked elements of varying sizes and, in distal rows decreasing in number and size. Radula formula 29 x 24.0. 24 in preserved specimen of 2.0 mm in length; innermost lateral teeth with enlarged base and cusp curved towards to base of tooth with denticles; marginal teeth elongated, slender, with finger-like projections on their distal part.
Reproductive system ( Figure 16 View FIGURE 16. A ): hermaphrodite duct connected to long and slightly convolute ampulla. Postampullary gonoduct short, that connects to oviduct and prostate. Prostate granular, divided into two parts; longer and less dense part proximal, and smaller and denser distal part; in juveniles prostate completely covers bursa copulatrix. Vas deferent elongate and not covoluted, that emerges from a common atrium with vagina. Vagina elongate, approximately of same width as deferent duct, opening into rounded bursa copulatrix. Bursa serially arranged, vaginal duct not convoluted and connecting to rounded seminal receptacle; very short uterine duct inserting, almost immediately, into female gland mass. In juveniles specimens, bursa copulatrix approximately of same size as seminal receptacle, while in sexually mature specimens bursa copulatrix three-four times larger in volume and length than seminal receptacle.
Biology. Egg mass composed by 2–3 rows, with red large eggs and a translucent material surrounding eggs individually in each capsule.
Remarks. Rostanga byga Er. Marcus, 1958a was described from Brazil and recorded three times (each record is based on one specimen) (Er. Marcus, 1958a; Ev. Marcus,1971; Padula et al., 2012). Subsequently, Muniaín and Valdés (2000) re-described the species based on the holotype and additional material from Argentina, but they could not analyze the reproductive system and radula of the holotype. They compared it to the highly similar Rostanga pulchra MacFarland, 1905 . However, after analyzing specimens of R. byga , we now question certain characteristics used by Muniaín and Valdés (2000) to separate these two species. Schrödl and Grau (2006), who analyzed Chilean specimens of R. pulchra , already had challenged some of these differences.
One of the distinguishing characteristics listed by Munían and Valdés (2000) is the presence or absence of white points that form a midline on the dorsum. This feature is not consistent—we identified specimens lacking white spots on the dorsum or with very faint white spots, probably due to the small size of the specimens. In some cases, there seem to be dark spots on the back, which was reported to be a characteristic of R. pulchra (Muniaín & Valdés, 2000) . Additionally, Muniaín and Valdés (2000) compared the number of perfoliations of the rhinophores between these two species and have concluded that R. pulchra has between nine and 12 perfoliation, whereas R. byga has 12 to 16 perfoliations. However, in the R. byga specimens analyzed in this study, nine perfoliations were found, hence this is not a consistent feature.
The differences concerning the radula are also questionable. Muniaín and Valdés (2000) considered that the cusp of the innermost lateral tooth is smaller and with more denticles in R. pulchra than in R. byga . However, our analysis of the micrographs presented in Schrödl and Grau (2006), Muniaín and Valdés (2000), and in the present study showed that the ranges of cusp lengths in R. pulchra and R. byga overlap (9.1 µm to 14.5 µm in R. pulchra ; 6.6 µm to 17.7 µm in R. byga ). Furthermore, in the specimens analyzed here, we observed that in the same radula there are significant differences in the length of the cusp of the innermost lateral tooth (ranges from 6.6 µm to 12.3 µm). We agree with Schrödl and Grau (2006) that it is difficult to visualize the number of denticles on the inner lateral tooth due to the position of the tooth in the radula. Thus, we will not estimate the number of denticles.
Muniaín and Valdés (2000) described R. pulchra as presenting a labial cuticle with regularly disposed elements, whereas R. byga presents a reduced labial cuticle with irregularly disposed elements. However, they do not provide an explanation for the difference, since in both cases the elements are disposed in rows. Interpreting the illustration presented in Muniaín and Valdés (2000: 4, 9, Figs. 2 View FIGURE 2. A – B E, 6E), we believe that the authors referred to size of the elements, in that uniformly sized elements correspond to R. pulchra and those with different-sized elements are R. byga . Based on this assumption, the specimens collected on the coast of Rio de Janeiro state are closer to the description given for R. byga ( Fig. 15 View FIGURE 15 A).
Our data corroborate the idea of Er. Marcus (1958a) and Schrödl and Grau (2006) that R. byga and R. pulchra are quite similar, and there is biogeographic evidence that assists the separation of the two species. Nonetheless, morphological studies of additional specimens of both species are needed to fully understand the intraspecific variation and differences between the two.


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |