
Megalothorax processus Panina & Potapov, 2022
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5188.4.6 |
|
publication LSID |
lsid:zoobank.org:pub:B35CE39E-9B1B-454D-A46A-08EBA6EC3CE5 |
|
DOI |
https://doi.org/10.5281/zenodo.7105224 |
|
persistent identifier |
https://treatment.plazi.org/id/0389879E-FFDA-CA43-34AA-FF16FC4AFB72 |
|
treatment provided by |
Plazi |
|
scientific name |
Megalothorax processus Panina & Potapov |
| status |
sp. nov. |
Megalothorax processus Panina & Potapov View in CoL , sp. nov.
Figs 18–35 View FIGURES 18–23 View FIGURES 24–26 View FIGURES 27–30 View FIGURES 31–35 , Table 1‒2 View TABLE 1 View TABLE 2
Type Material. Holotype, Russia (European part), Nenets Autonomous Okrug, Yugorsky Peninsula, surroundings of Amderma [69075′ N, 610 67′ E], meadow with dryad, 15– 16.07.2018, leg. M. Bizin and B. Efeikin. 10 paratypes, in same location. The holotype and 8 paratypes deposited in the collection of the Department of Zoology & Ecology, Moscow State Pedagogical University. 2 paratypes deposited in SMNG.
Other material. Russia (Asian part), Republic of Altai, Chuysky tract, Kosh-Agachsky district , at the exit from the Chuysky basin [50 ° 10′ N, 88 ° 34′ E], floodplain forest (larch with willow undergrowth), 06.10.2020 ( Fig. 37 View FIGURES 36–37 ); GoogleMaps Altai Republic, Chuisky tract, Ongudai district , Seminsky pass [50 ° 99′ N, 85 ° 65′ E], meadow with Betula nana in brook valley, mosses on a bump, 04.10.2020. All leg. M. Potapov and N. Kuznetsova. Novaya Zemlya, GoogleMaps South Island , Bezymyannaya River Valley [72050′ N, 53045′ E], willow-herb thickets, 20– 26.07.2017, leg. V. Spitsin; Nenets Autonomous Okrug, Pakhanchenskaya Guba [68029′ N, 570 12′ E], forb meadow on slope, 26.07.– 05.08.2015, leg. O. Makarova and M. Bizin. All kept in MSPU GoogleMaps .
Description. General aspect. Habitus and segmentation typical of the genus. Body length up to 0.35 mm. Specimens whitish in alcohol. Body chaetotaxy sparse including chaetae, s-chaetae, trichobothria, neosminthuroid chaetae, wax-rods and inner sensilla within sensory fields 2–6. Chaetae ordinary on body, without any remarkable development.
Integument. As in M. roseus sp. nov. Connection of channels with linea ventralis circular on the head.
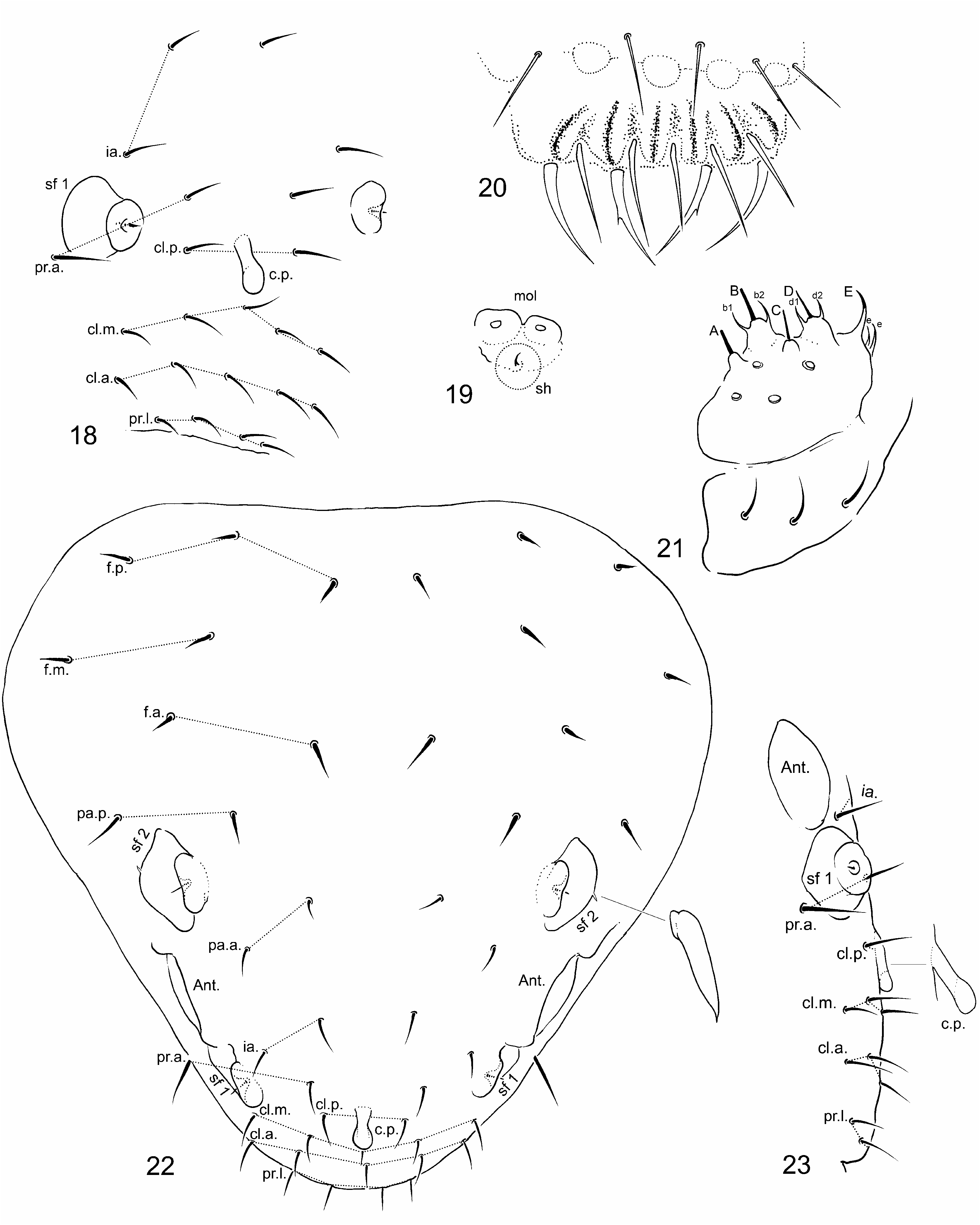
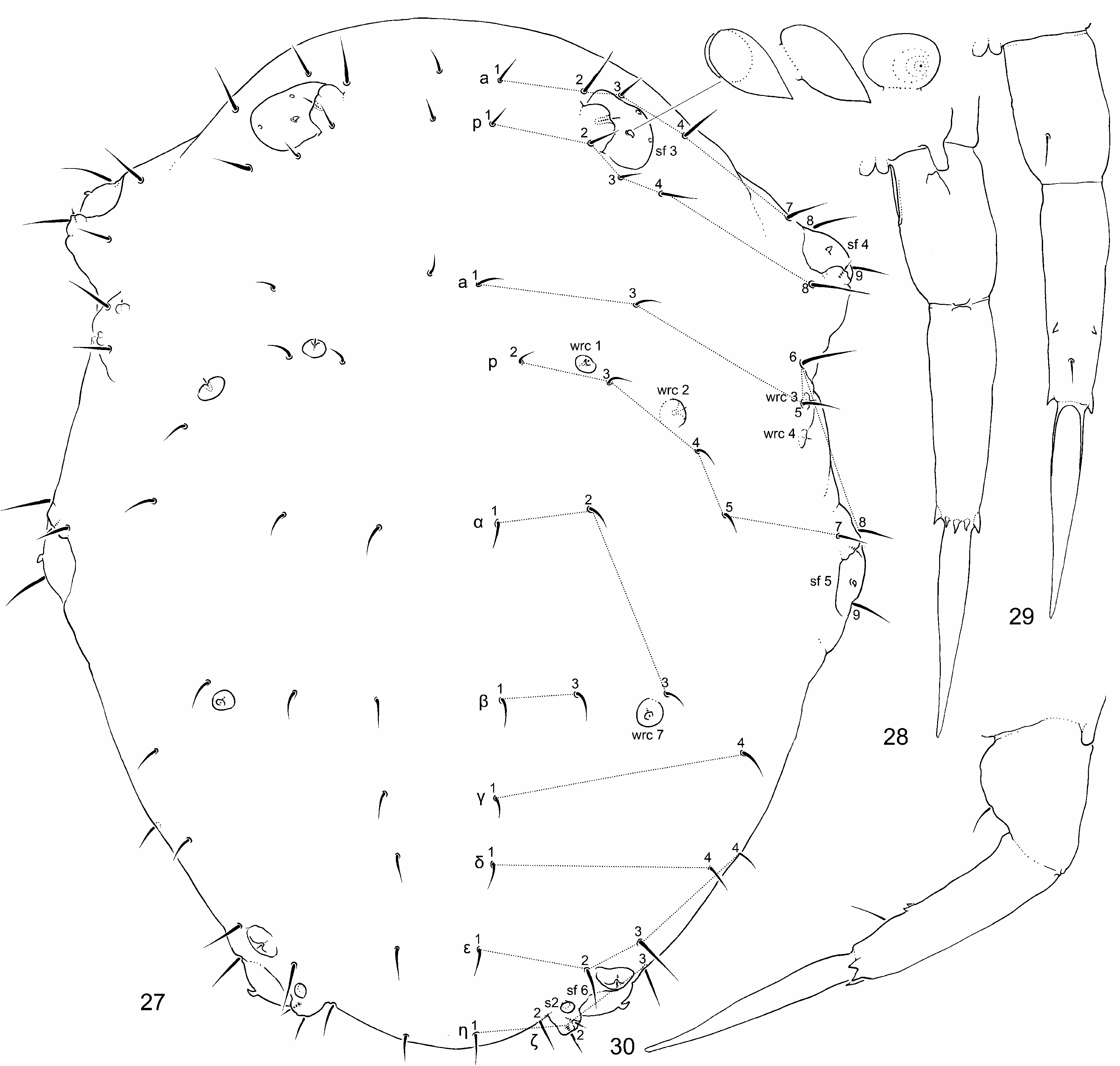
Sensory fields and wax rods ( Figs 22 View FIGURES 18–23 , 27 View FIGURES 27–30 , 31 View FIGURES 31–35 ). Sensory field sf 1 without inner sensilla (s) and with one wrcchaeta. Sf 2 with one inner s and one wrc-chaeta (wrc). Sf 3 with three s and one wrc. Sf 4 and sf 5 with two s and one wrc. Sf 6 with one s and two wrc. Each s of sf 2–6 broad flame-shaped ( Figs 27 View FIGURES 27–30 , 31 View FIGURES 31–35 ). A total of 14+14 wrc (2 + 2 on head, 12 + 12 on body), including free 7+7 wrc (not associated with sensory fields, notated as wrc 1–wrc 7).
Mouthparts. Labrum typical of the genus ( Fig. 20 View FIGURES 18–23 ). Chaetae a1 and a2 not forked, with one or two teeth. Labium with 4+4 proximal chaetae ( Fig. 21 View FIGURES 18–23 ). Basomedian field with 3+3 chaetae. Basolateral field with 1 + 0 chaetae (one tubercle and no ventral chaeta). Labial palp as in Fig. 21 View FIGURES 18–23 , as common for the genus (A,B,C,D,E, b1, b2, d1,d 2, 2e, H, h1, h2). Oral fold and maxillary outer lobe typical of the genus, with one sublobal hair (sh). Maxillary head without strong modification.
Head chaetotaxy. Forehead chaetotaxy as on Figs 18, 22, 23 View FIGURES 18–23 . Clypeo-labral formula: 2, 5, 5, 4/ 5, 5, 4 ( Fig. 18 View FIGURES 18–23 ). Chaetae a0 absent, replaced by a long cuticular process extended at the tip ( Figs 18, 22, 23 View FIGURES 18–23 ). Dorsal posterior area with 18 lanceolate chaetae ( Fig. 22 View FIGURES 18–23 ). Ventral side with three pairs of postlabial chaetae. Trend for posterior chaetae to be longer and stronger than anterior chaetae.
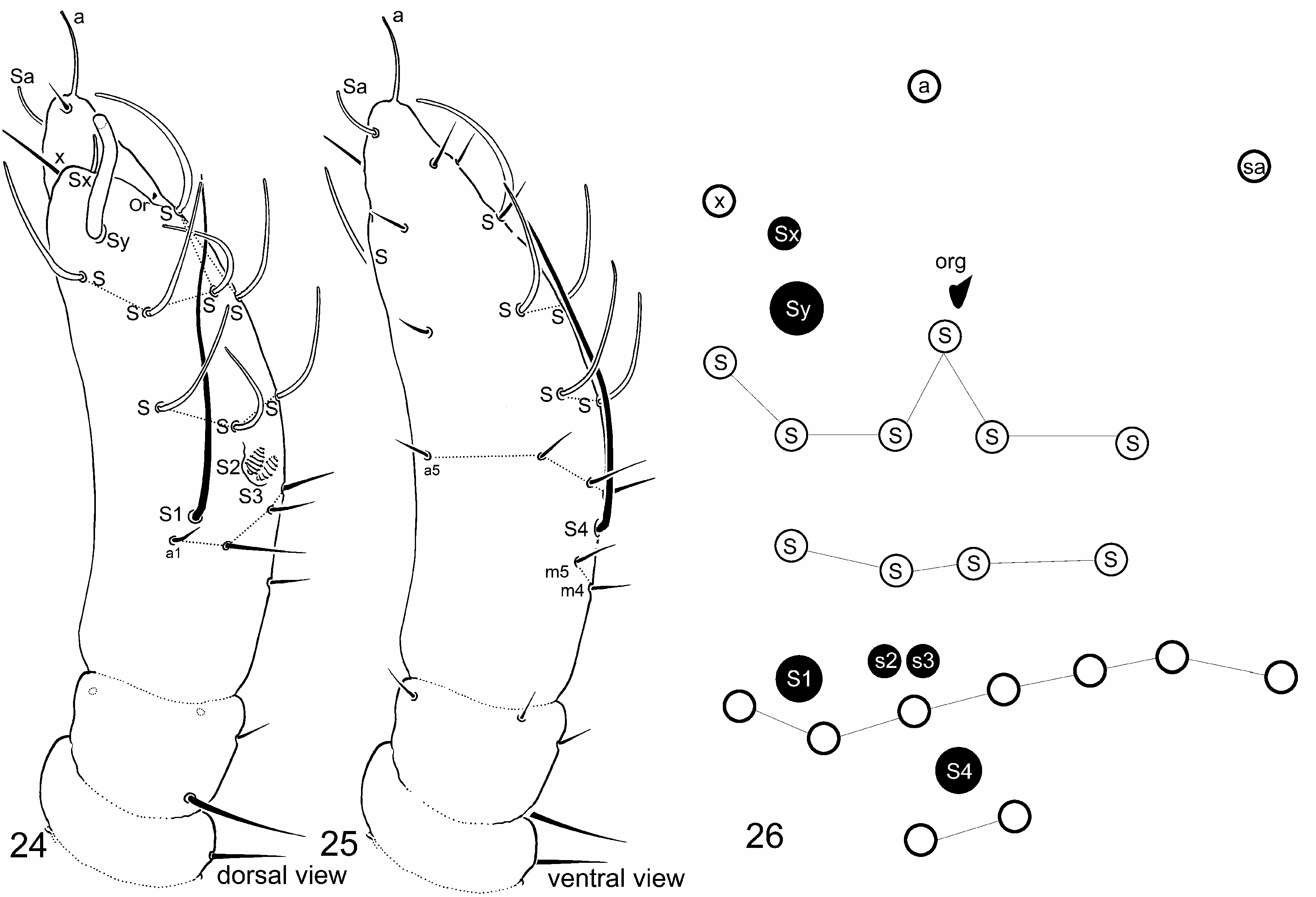
Antennal chaetotaxy ( Figs 24–25 View FIGURES 24–26 ). Pattern diagram in Fig. 26 View FIGURES 24–26 . Ant. I and II with one and four chaetae, respectively. Dorsal chaeta on the Ant. II bigger than others. Ant. III with 8–9 chaetae, two long S-chaetae (S1 and S4). Striations of Ant III sensory organ short sensilla (S2 and S3) distinguishable in light microscopy. Ant. IV with seven chaetae (including X-chaeta) and ten S-chaetae. Sensory organ with Sx, Sy, Or, a, Sa. Organite (Or) of Ant IV short, seems apically flared. Summary on antennal chaetotaxy provided in Table 1 View TABLE 1 .
Body chaetotaxy. Th. II with 12+12 chaetae, 1 + 1 tubular and curved s1-sensilla ( Figs 27 View FIGURES 27–30 , 31 View FIGURES 31–35 ). Th. III with 11 + 11 chaetae, 6 + 6 free wax-rods (wrc1–6). Chaetae p4 not close to wrc2 ( Figs 27 View FIGURES 27–30 , 31 View FIGURES 31–35 ). Chaeta a5 slightly shorter than chaeta a6. Abd I–V terga with 17 + 17 ordinary chaetae, 1 + 1 free wax-rods, 1 + 1 globular sensillum s2. Globular sensillum s3 absent ( Figs 27 View FIGURES 27–30 ). Chaetae of body subequal, slightly thickened.
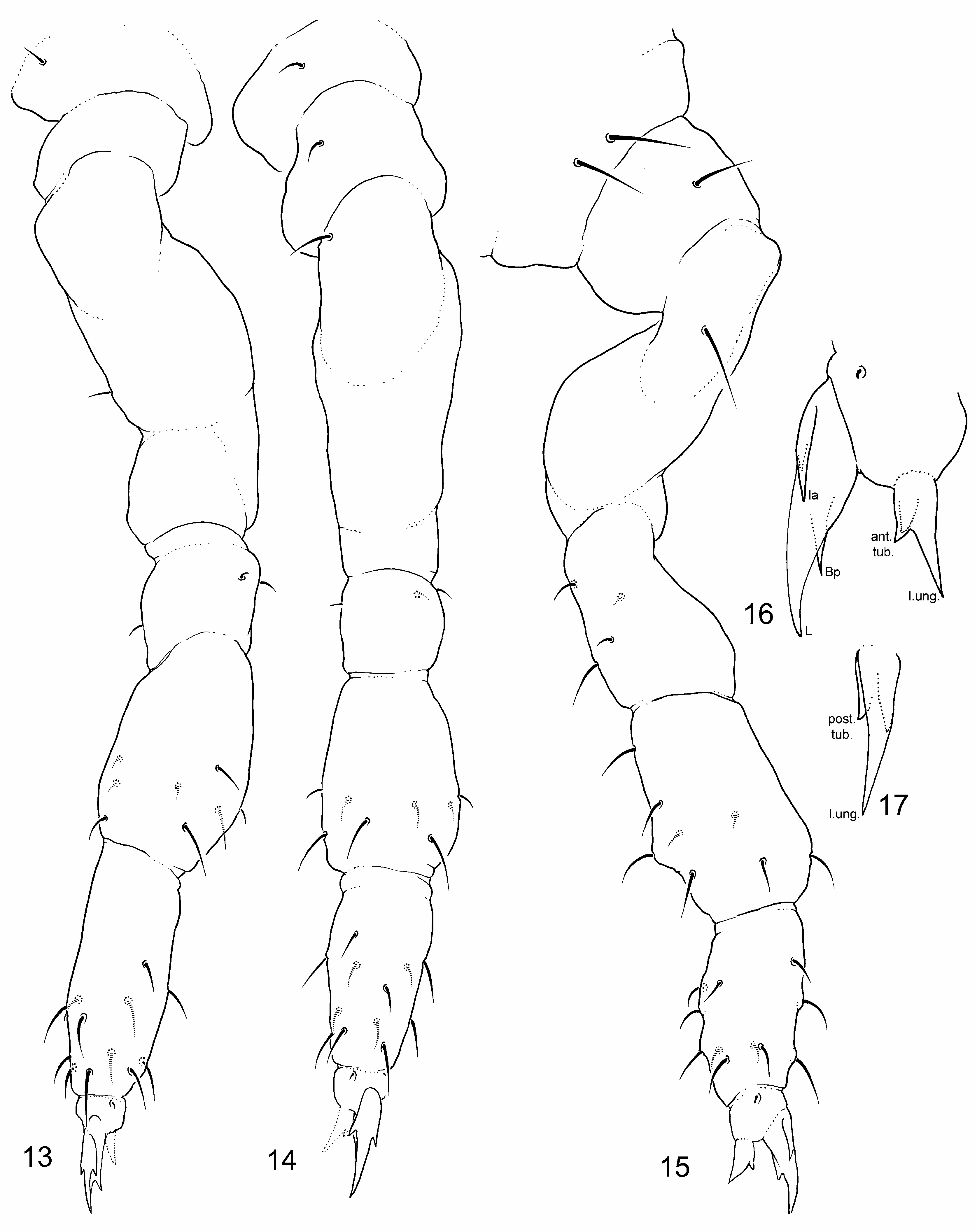
Legs chaetotaxy. Leg chaetotaxy consist of ordinary chaetae of variable size, about as in Figs 13–15 View FIGURES 13–17 . Chaetotaxy leg typical of the genus. Number of chaetae for each segment is summed up in Table 2 View TABLE 2 .
Claws. Claw III bulkier than claw I and II ( Figs 32–34 View FIGURES 31–35 ). Claws subequal in unguis length (with a trend as unguis I>unguis II> unguis III). Unguis basal and posterior auxiliary lamellae (la, lp and Bp) well developed ( Figs 32–34 View FIGURES 31–35 ). Unguiculus about 0.6 as long as unguis.
Abd. IV sternum and furca. Abd. IV sternum with 2 + 2 neosminthuroid chaetae and at least 2+2 chaetae (the observations are uncertain). Manubrium with 2 + 2 posterior chaetae and 1 + 1 pegs with convex tip articulated with a corresponding concavity of the dens ( Fig. 28 View FIGURES 27–30 ). Dens as in M. roseus sp. nov. ( Figs 28–30 View FIGURES 27–30 ). Mucro narrowing in the distal 2/5. Edges are entirely smooth with one notch. Chaetotaxy of Abd. V and VI not studied, looks generally as for the genus.
Tenaculum and ventral tube. Tenaculum with 3 + 3 hook-like teeth ( Fig. 35 View FIGURES 31–35 ). Ventral tube bulky with two apical pairs of chaetae.
Males not found.
Name derivation. The name reflects the presence of a process on the forehead.
Discussion. Megalothorax processus sp. nov. belongs to the minimus -group ( Schneider & D’Haese 2013). Specific cuticular process is usually well visible and easy defines this species. Formally, M. potapovi and M. sanctistephani also have the front head process, albeit fold- and coffee bean-shaped, respectively (vs. long and extended at the tip in M. processus sp. nov.). M. potapovi also differs in number of chaetae on the forehead, including chaeta a0 (present in M. potapovi vs. absent in M. processus sp. nov.) and body (more chaetae in M. potapovi ). M. sanctistephani lacks chaeta X on Ant.IV (present in M. processus sp. nov.) and has 2 sublobal hairs on maxilary outer lobe (vs. 1 in M. processus sp. nov.). M. processus sp. nov., M. roseus sp. nov. and M. laevis share the reduced chaetotaxy. Both new species differ from M. laevis by the presence of process, the absence (vs. presence) of sensilla s3 on body and 3 + 3 (vs. 4+4) teeth on tenaculum. The new species differs from widely distributed M. willemi and M. minimus by many characters, including the process and smaller flame-shaped inner sensilla. M. processus sp. nov. differs from M. roseus sp. nov. with shape of the process, chaetotaxy of forehead, number of sublobal hairs, and shape of inner sensilla in sensory field.
The four species ( M. sanctistephani , M. potapovi , M. roseus sp. nov., M. processus sp. nov.) belong to minimus - group (in spite of doubts on M. roseus sp. nov.) Their “nose”, however, is unlikely homologous if considering its shape and location. The independent appearance of this process probably indicates its importance to these species. The function of small unpaired “nose” is, presumably, delicate unlike, e.g., in stiff epistome in Acari (which protects mouth parts). In Collembola with similar position of head ( Symphypleona ), specific equipment on head front appear in sexually dimorphic taxa only (e.g., "nasal" organ in Nasosminthurus Stach ) which cannot be associated to mentioned congeners of Megalothorax at all because of parthenogenesis.
Distribution and ecology. Megalothorax processus sp. nov. is recorded from the arctic deserts to the mountains of Southern Siberia. It occurs in meadows, floodplain forests, and thickets along rivers. It is probably noted as Megalothorax sp. 2 in the East-European tundra ( Babenko et al. 2017).


| SMNG |
Senckenberg Museum fuer Naturkunde Goerlitz |
| V |
Royal British Columbia Museum - Herbarium |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |