
Pelodytes hespericus, Díaz-Rodríguez, Jesús, Gehara, Marcelo, Márquez, Rafael, Vences, Miguel, Gonçalves, Helena, Sequeira, Fernando, Martínez-Solano, Iñigo & Tejedo, Miguel, 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4243.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CE502CF7-6F19-43A2-AD79-15DBE777E28B |
|
DOI |
https://doi.org/10.5281/zenodo.6016578 |
|
persistent identifier |
https://treatment.plazi.org/id/038A7153-7073-BC6C-95A2-FB8CFCE65EC2 |
|
treatment provided by |
Plazi |
|
scientific name |
Pelodytes hespericus |
| status |
sp. nov. |
Pelodytes hespericus View in CoL sp. nov.
( Fig. 12 View FIGURE 12 )
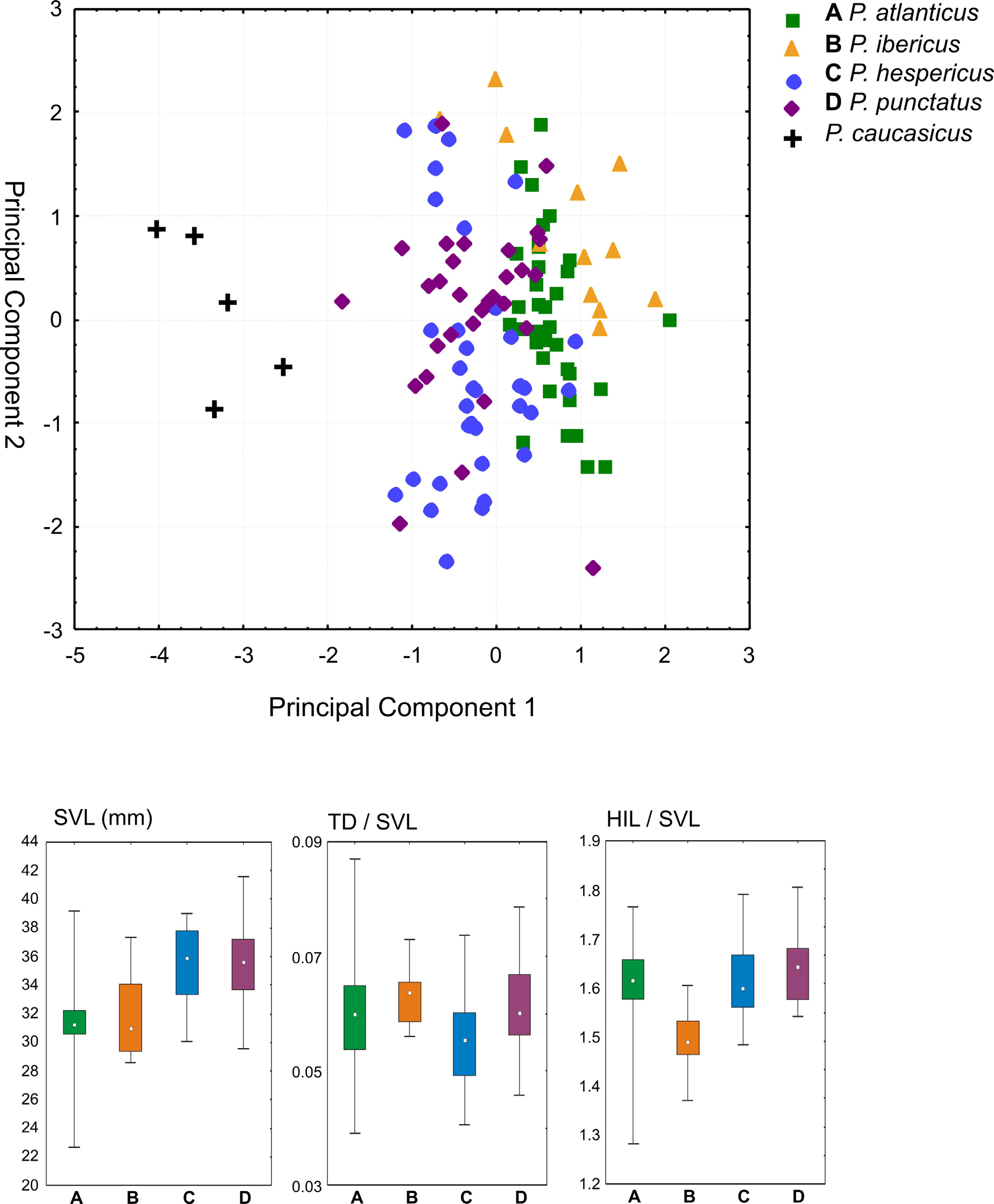
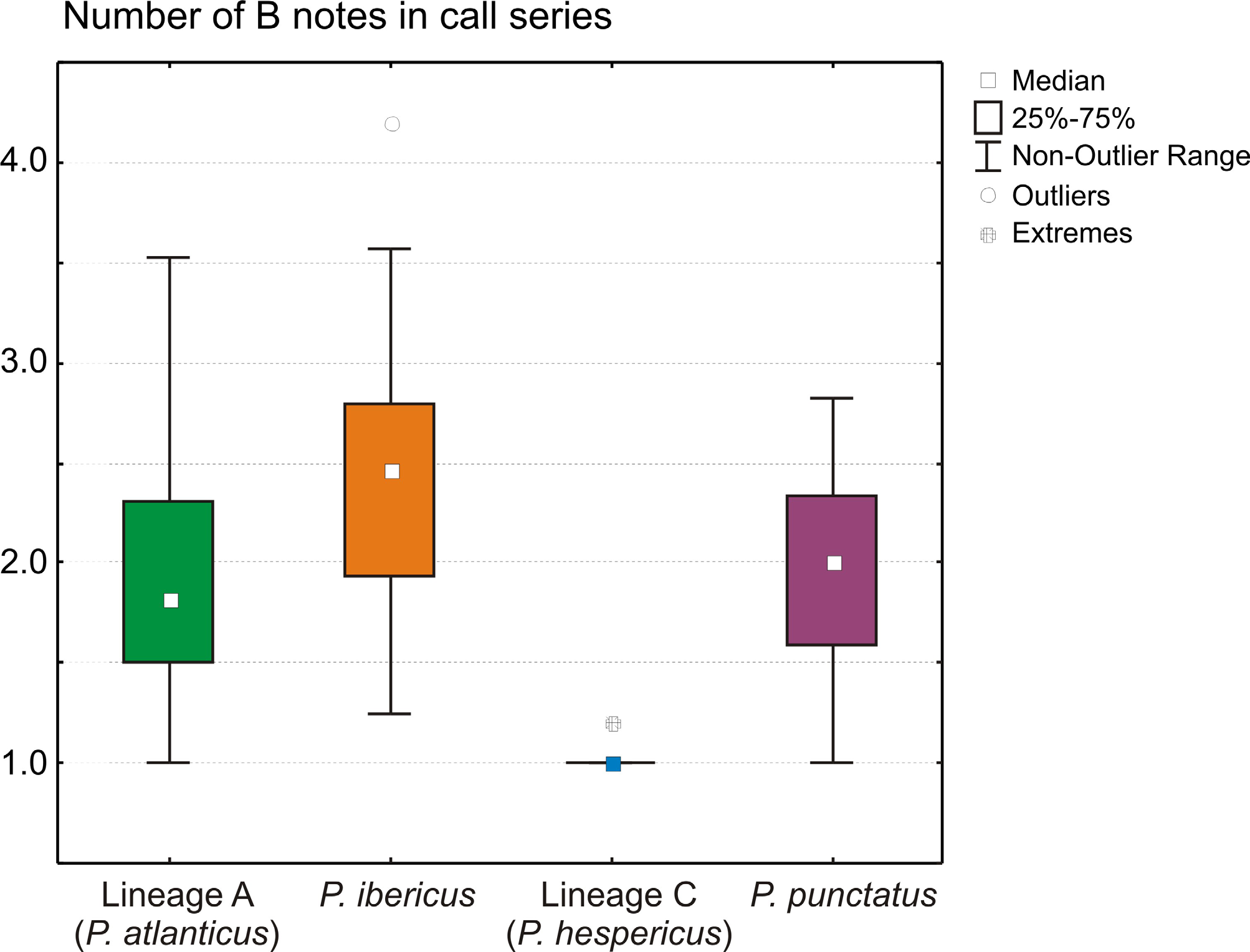
Identity and diagnosis. Populations of this species have been considered as P. punctatus by Sánchez-Herráiz et al. (2000), and as lineage C by Díaz-Rodríguez et al. (2015). Assigned to the genus Pelodytes based on high morphological similarity to P. punctatus , the type species of the genus; vertical pupil; network of dark lines in the skin of tadpoles; tadpoles with sinistral spiraculum; and molecular phylogenetic relationships. Of similar size as P. punctatus ; mean SVL 35.3 mm (maximum 39.0 mm) in males, 39.8 mm ( 43.3 mm) in females. Morphologically similar to the other three western Pelodytes species, but distinguished from all of them by no repetitions of B notes in advertisement calls according to the recordings analyzed herein ( Fig. 7 View FIGURE 7 ), from P. ibericus by longer limbs and larger body size, and from P. atlanticus sp. nov. by larger body size ( Fig. 6 View FIGURE 6 ; Table 3 View TABLE 3 ). Furthermore, it is distinguished from all other Pelodytes species by concordant differences in mtDNA and nDNA sequences.
Holotype. EBD 34505 (JDR 2012-1), adult male collected by Jesús Díaz-Rodríguez on 9 April 2012 at Artunido ( Casas de Carrasco ), Sierra de Segura, Jaén province, Spain. Geo. coord: 38.119349, -2.691951.
Paratypes. EBD 34506 (JDR 2012-2), adult male, and EBD 34507 (JDR 2012-3), adult female, collected on 8 April 2012 by same collector and at same locality as holotype. Individuals deposited in Doñana Biological Station with numbers EBD 28169, 28170 , 28171, 28172, 28173, 28174, 28175 collected from Soria ; EBD 20440 collected from Huesca , EBD 33728 and EBD 26828 collected from Almería; and specimens deposited in National Museum of Natural Sciences with the numbers MNCN 667, MNCN 669, MNCN 670, MNCN 672, MNCN 673, MNCN 684, MNCN 686, MNCN 2720, MNCN 2721, MNCN 2722, MNCN 2723, MNCN 2724, MNCN 2725 About MNCN collected from Valencia ; MNCN 689 About MNCN collected from Ciempozuelos ( Madrid) ; MNCN 25023, MNCN 25024, MNCN 25025, MNCN 25026, MNCN 25027, MNCN 25028, MNCN 25029, MNCN 25032, MNCN 25033, MNCN 25034, MNCN 25035, MNCN 25036 About MNCN collected from La Rioja ; MNCN 41414, MNCN 41416 collected from Masa (Burgos) and MNCN 24489, MNCN 24490 About MNCN from Merindad del Río Ubierna ( Burgos) ; MNCN 25020 from Teruel, MNCN 25021, MNCN 25037 from Zaragoza, MNCN 41697 About MNCN from Brea de Tajo ( Madrid) , MNCN 41698 About MNCN from Corpa ( Madrid) , MNCN 377 from Almodóvar del Campo (Ciudad Real); MNCN 25022 collected from Palencia and MNCN 20783 collected from Castellón.
Etymology. The specific name is a latinized adjective derived from the name Hesperides ( Ἑσπερίδες), Nymphs of the fading sun and daughters of the Titan Atlas , in allusion to an Iberian mythological garden. The Hesperian Massif also takes its name from the same etymology, meaning “western” in ancient greek, referring to its position in the Mediterranean sea.
Description of the holotype. Adult male in excellent state of preservation. Some tissue removed ventrally from right thigh for molecular analysis. Measurements: SVL, 36.6; HW, 12.7; HL, 13.3; TD, 2.7; ED, 4.0; END, 4.3; NSD, 3.0; NND, 3.6; HAL, 10.6; FORL, 23.2; HIL, 61.6; FOL, 18.4; FOTL, 28.5; TIL, 18.4. Body relatively slender; head dorsally flattened, slightly longer than wide, wider than body; snout slightly pointed in dorsal view, rounded in lateral view; nostrils directed dorsolaterally, not protuberant, nearer to tip of snout than to eye; canthus rostralis indistinct, straight; loreal region very slightly concave, almost flat; eye moderately large, notably protuberant dorsally; interocular distance slightly smaller than horizontal eye diameter, larger than internarial and slightly smaller than eye-nostril distance; pupil vertical; tympanum externally visible, distinct, oval, wider vertically than horizontally, its horizontal diameter is 68% of maximum eye diameter, positioned near corner of mouth; supratympanic fold moderately distinct in its anterodorsal part running from posterior corner of eye to posterior corner of tympanum, indistinct in its posteroventral part running from posterior corner of tympanum to insertion of forelimb; tongue free posteriorly, of truncate shape, vomerine teeth distinct, in two patches between and at the same level as choanae; choanae moderately sized, ovoid. Premaxillary and maxillary teeth present. Arms slender, webbing almost absent, very small rudiments recognizable; comparative finger length I<II<IV<III, fingers with dermal fringes (poorly recognizable on small fingers due to fixation). No enlarged finger disks. One large, rounded subarticular tubercle present proximally on each finger; three flattened metacarpal tubercles; inner and outer metacarpal tubercles of similar size and distinctly larger than central tubercle. Black nuptial pads on inner sides of first and second fingers, as one elongate patch on forearm, a second slightly smaller elongate patch on arm, and a small agglomeration on both sides of chest near axilla. Hindlimbs slender; tibiotarsal articulation reaches between eye and nostril when adpressed along body; inner metatarsal tubercle distinct but relatively small, outer metatarsal tubercle absent; proximal subarticular tubercles indistinct, remaining subarticular tubercles absent. Comparative toe length I<II<V<III<IV; third toe distinctly longer than fifth toe; toes with conspicuous dermal fringes to tip. Webbing between toes weakly expressed but clearly present and more than merely an extension of dermal fringes. Webbing formula difficult to determine due to absence of subarticular tubercles and presence of dermal fringes, but webbing almost complete between toes I and II, between second and third phalangeal joint of toe II between toes II and III. Skin on the dorsal surface granular, with distinct warts; skin of throat, chest, and venter smooth except granular on posteriormost part of belly; ventral surfaces of thighs and arms granular. Especially those warts that coincide with dark greeninsh spots on dorsum of slightly elongated shape and arranged in longitudinal rows. The anterodorsal part of the supratympanic fold is continued into the inguinal region as a distinct row of small tubercles and ridges, partly marked by greenish dots.
Colour of the holotype. The skin colour is between dark green to olive drab with light olive bands on the back and legs. The back is dotted with a pattern of small oval and linear prominences of olive green. On the lateral side, a clear band is more or less visible, from the tympanum to the insertion of posterior limbs. The ventral area, which is smooth or slightly rough, has a light colour from pale green to yellow tones. Colour in life is similar, with light greenish spots on the back, instead of drab colors, and a design of two crossed bands clear without dots ( Fig. 12 View FIGURE 12 ).
Variation. In the paratypes EBD 34506 and EBD 34507, the foot webbing is much less distinctly expressed.
Distribution. Pelodytes hespericus is distributed in the Iberian Peninsula and as far as known, restricted to Spain. Its range extends mainly in areas of low and mid-altitude mountains, from the sea level to 1,900 m in La Rioja ( Zaldívar 2004), almost 2,000 m in Sierra Seca ( Granada) and maybe higher at the locality of Castril, Granada ( Fernández-Cardenete et al. 2000), but additional confirmation is required. It occupies most of the eastern and central parts of Spain (Lacomba & Sancho 1999; Barbadillo 2002b). The western range limits are in the southeast of Madrid ( Martínez-Solano & García-París 2001; Martínez-Solano 2006), in the provinces of León and Zamora, and in the Montes de Toledo in Toledo and Ciudad Real. Southwards the species reaches the northern parts of Jaén and Granada provinces ( Barbadillo 2002b; Ceacero et al. 2007) and coastal parts of Almería (González- Miras & Nevado 2008; Díaz-Rodríguez et al. 2015).
Natural history. Lizana et al. (1986) and Esteban et al. (2004) provided notes on the natural history of this species. Pelodytes hespericus is a generalist that tolerates a wide range of climatic conditions and diverse substrates. However, it seems to avoid siliceous and some types of acid soils (Sánchez-Herráiz 2004). It can be found in semi-arid steppes in the Ebro basin and the southeast, as well as in temperate forests of beech and oak, but usually avoids the densest forest formations. It is common in abandoned quarries and cattle ponds and tolerates high levels of salinity. Specimens can reach eight years (males) and 10 years (females) of age ( Esteban et al. 2004; Sánchez-Herráiz 2004).
Advertisement call and reproductive behavior. As in the other western Pelodytes lineages, the call consists of two types of notes ( A and B), nonetheless emitted just once time. In the recordings previously studied by Sánchez-Herráiz (2004) analyzed by us, and in numerous other individuals heard in the field, one call typically consists of one A note followed by a single B note ( Fig. 7 View FIGURE 7 ). Reciprocal calls of females have been reported (Sánchez-Herráiz 2004). Calling specimens are found at the edge or within water, with body temperatures between 10 °C –17.5°C (Sánchez-Herráiz 2004; Esteban et al. 2002). Call intensity and spectral frequency were not found to be related to the size or age of the male ( Esteban et al. 2002).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.