
Moroxylon, SELMEIER, 1933
|
publication ID |
https://doi.org/ 10.5070/P9401462457 |
|
persistent identifier |
https://treatment.plazi.org/id/038AF505-A327-963A-5573-F916FD5F9D73 |
|
treatment provided by |
Felipe |
|
scientific name |
Moroxylon |
| status |
|
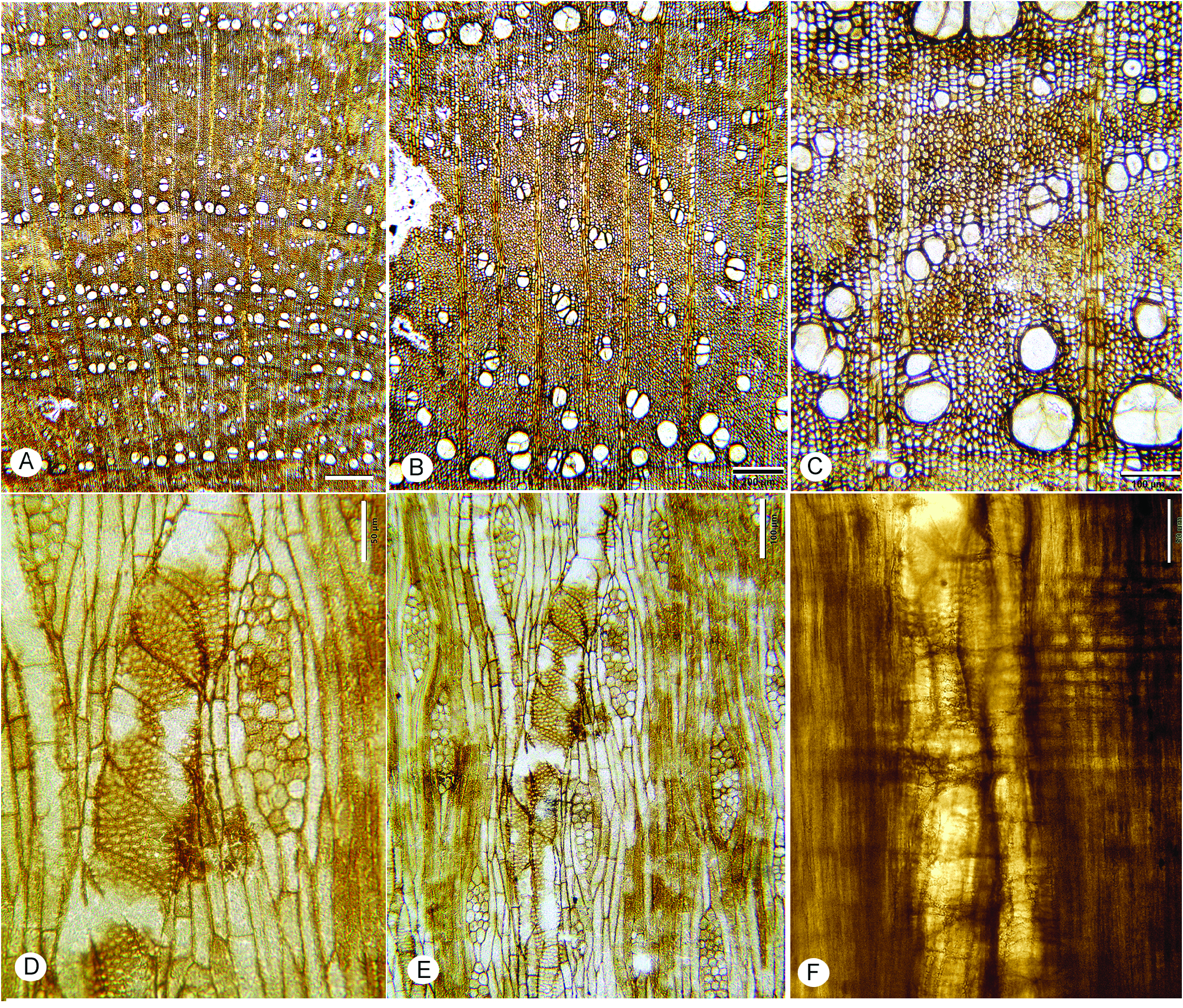
FIG. 12A–F View Figure 12
Description— Growth rings present, marked by radially narrow fibers, and marginal axial parenchyma ( Fig. 12A, B View Figure 12 ).
Wood ring-porous; earlywood zone narrow with 1–2 rows of earlywood vessels that are rounded in outline ( Fig. 12A–C View Figure 12 ); mean tangential diameter of earlywood vessels 91 µm ( SD =16), range 67–132 µm; latewood vessels often in a diagonal arrangement, solitary and in radial multiples ( Fig. 12B View Figure 12 ), vessel clusters uncommon. Perforation plates simple ( Fig. 12D‒F View Figure 12 ); intervessel pits alternate, polygonal in outline ( Fig. 12D View Figure 12 ), horizontal di- ameter 8–11 µm. Vessel-ray parenchyma pits similar in size to intervessel pits ( Fig. 12F View Figure 12 ). Vessel elements short, typically less than 200 µm. Thin-walled tyloses. Helical thickenings not observed.
Fibers thin- to thick-walled, mostly non-septate, occasionally septate ( Fig. 14D, E View Figure 14 ), pitting not observed. Vasicentric/vascular tracheids absent.
Axial parenchyma marginal and confluent ( Fig. 12A– C View Figure 12 ); strands of 2–4 cells ( Fig. 14D, E View Figure 14 ).
Rays 1–3 (-4) seriate; uniseriate rays rare, average multiseriate ray height 263 ( SD =65) µm, 166–432 µm. Rays homocellular, composed of procumbent cells, and heterocellular with 1-2 marginal rows of square/upright cells ( Fig. 12D–F View Figure 12 ), rarely more, 4–6 per mm.
Storied structure and oil/mucilage cells not observed.
Specimen— UF 278-84899, estimated maximum diameter 4 cm.
Occurrence— Dietz Hill ( UF 278).
Comments— The intervessel pits and most of the vessel-ray parenchyma pits have a similar size, but we found it difficult to determine whether the vessel-ray pa- renchyma pits did or did not have reduced borders. The occurrence of tyloses suggests that they have reduced borders ( Chattaway 1949, Bonsen and Kucera 1990, Wheeler et al. 2007).
Comparisons with modern woods— We searched InsideWood for ring-porous woods (3p) with vessels solitary and in short radial multiples (9a 10a), latewood with a diagonal pattern (7p), simple perforation plates (13p), alternate intervessel pitting that is not minute (22p 24a), vessel-ray parenchyma pitting both similar to intervessel pits (30p) and with reduced borders to apparently simple (31p), non-septate fibers with simple pits (61p 66p), confluent axial parenchyma (83p), rays not exclusively uniseriate or>10-seriate (96a 99a), rays not comprised of all upright cells (105a) and not markedly heterocellular (108a), storied structure absent (118a 120a), oil/mucilage cells absent (124–126a), and radial canals absent (130a).
This search with 30p and 31p gave two exact matches: Celtis australis L. (1753) and Broussonetia papyrifera ( L.) l'Hér. ex Vent (1799). If only 30p is used, the results also include Catalpa bignonioides Walter. , Ca. speciosa Teas (1875; Bignoniaceae ), Celtis tournefortii Lam. (1797; Cannabaceae ), Ehretia acuminata R.Br. (1810; Boraginaceae Juss.,1789 ), Gymnocladus dioicus ( L.) K.Koch (1869), Platyosprion platycarpum Maxim. (1877; Fabaceae ), Melia azedarach L. (1753; Meliaceae ), Phellodendron chinense C.K. Schneid. (1907) , Ptelea trifoliata L. (1753; Rutaceae Juss., 1789 ). If only 31p is used, the results also include Rhus chinensis Mill. (1768; Anacardiaceae ) and R. virens Lindh. ex A. Gray (1850) , Ce. biondi Pamp. (1910; Cannabaceae ), Ce. bungeana Blume (1852), Ce. jessoensis Koidz., Morus alba ( Moraceae ), Mo. cathayana Hemsl. (Forbes and Hemsley, 1894) , and Mo. serrata Roxb. (1832) . It seems most probable that this wood belongs to the Urticales group.
Narrow earlywood zones as seen in this wood are fairly common in Celtis as shown by a review of images in the Chinese Wood Atlas ( Itoh et al. 2022), e.g., Ce. biondi, Ce. bungeana, Ce. sinensis Pers. (1805) , Ce. tetrandra Roxb. (1832), Ce. vandervoetiana C. K. Schneid. (Sargent 1917). InsideWood’s image collection shows this feature also occurs in Ce. australis . Ce. laevigata Willd. (1814) , and Ce. occidentalis L. (1753). However, these species usually have obvious vessel clusters in latewood, unlike this fossil, and wider rays, usually with obvious sheath cells. Also, prismatic crystals occur in all Celtis woods described to date. Septate fibers do not occur in the Cannabaceae .
Earlywood zones with 1–2 rows of wide vessels also occur in some samples of ring-porous Morus , e.g., Mo. alba , Mo. australis Poiret (1796) , Mo. bombycis Koidz. (1915a) , Mo. cathayana , Mo. kagayamae Koidz. (1915b) (FFPRI’s Wood Database, https://db.ffpri.go.jp/Wood- DB/index-E.html). Vessel clusters and prismatic crystals are not constant features of Morus . Consequently, we suggest this wood’s affinities are most likely with the Moraceae .
Comparison with fossil woods— We used InsideWood’s fossil wood menu to search the fossil wood database and used the features: wood ring-porous (3p), vessels not in dendritic arrangement or exclusively solitary or commonly in radial multiples of four or more (8–10a); simple perforation plates (13p); intervessel pits alternate and not minute (22p, 24a); fibers with simple pits (61p); confluent axial parenchyma (83p), rays be- tween 2–10 cells wide (96a, 99a) and not composed of all upright/square cells or markedly heterocellular (105a, 108a); tile cells, storied structure, radial canals, and oil/ mucilage cells all absent (111a, 303a, 120a, 130a, 304a). Ignoring the fossil wood descriptions with a majority of features coded as unknown (?), a search allowing no mismatches returned two Cannabaceae , one Moraceae , one legume, one Rutaceae , and an Ulmaceae . The Cannabaceae , the legume, the Rutaceae , and the Ulmaceae are not good matches as they commonly have vessels in clusters and vessels in wavy tangential bands.
Moroxylon Selmeier (1993) is diagnosed as having latewood vessels commonly in clusters and in ulmiform tangential bands. However, vessel clusters do not look common in Selmeier’s figures 3, 4, and 13 and the ves- sel arrangement appears as much diagonal/oblique as tangential. Moroxylon sturmii Selmeier (1993) differs from this Dietz Hill wood in having an earlywood zone more than 1–2 vessels deep, occasional sheath cells and prismatic crystals in upright/square ray cells ( Table 3). We think that the similarities between them are such that we are referring this wood to cf. Moroxylon .
| SD |
San Diego Natural History Museum |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| K |
Royal Botanic Gardens |
| C |
University of Copenhagen |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |