
Magnolia hansnooteboomii, Wheeler & Manchester & Baas, 2023
|
publication ID |
https://doi.org/ 10.5070/P9401462457 |
|
persistent identifier |
https://treatment.plazi.org/id/038AF505-A339-962B-57D7-FCD9FDBF9E6E |
|
treatment provided by |
Felipe |
|
scientific name |
Magnolia hansnooteboomii |
| status |
sp. nov. |
MAGNOLIA HANSNOOTEBOOMII SP. NOV.
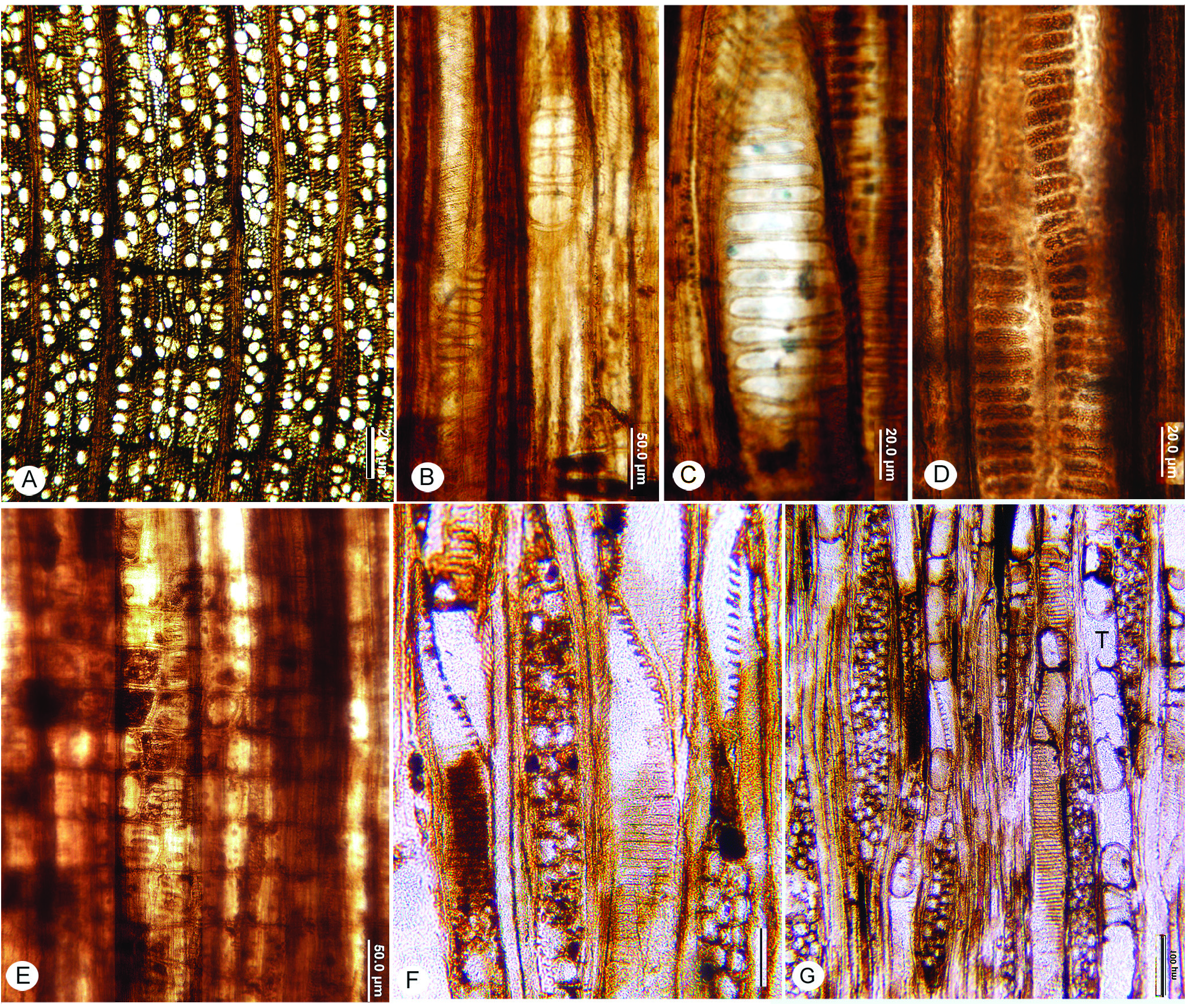
FIG. 4A–G View Figure 4
Diagnosis— Wood diffuse-porous. Vessels solitary and in radial multiples; vessels very narrow to narrow. Perforation plates exclusively scalariform with fewer than 20 bars; intervessel pits predominantly scalariform. Vessel-ray parenchyma pits horizontally elongate. Fine helical thickenings present throughout the body of each vessel element. Thin-walled tyloses present.
Fibers non-septate.
Axial parenchyma marginal.
Rays mostly 3–4 cells wide, uniseriate rays rare. Oil/ mucilage cells absent.
Holotype — UF 278-84871, estimated maximum diameter 7 cm
Etymology— In recognition of the late Hans Peter Nooteboom (1934–2022) and his many years of study of the Magnoliaceae .
Description— Growth rings present, marked by radially narrow fibers and marginal parenchyma.
Wood diffuse-porous; vessels solitary and predominantly in radial multiples of 2–4 ( Fig. 4A View Figure 4 ); mean tangential diameter 45 (9) µm, 28–68 µm; 75–100 per mm 2. Perforation plates exclusively scalariform with 4–11–17 bars (n=23) ( Fig. 4B, C, F View Figure 4 ); intervessel pits predominantly scalariform ( Fig. 4D View Figure 4 ). Vessel-ray parenchyma pits horizontally elongate and throughout the ray ( Fig. 4E View Figure 4 ). Fine helical thickenings present throughout the body of the vessel element ( Fig. 4B, C View Figure 4 ). Vessel element lengths average 609 (92) µm, range 407–746 (n=24). Thin-walled tyloses present ( Fig. 4G View Figure 4 ).
Fibers non-septate.
Axial parenchyma marginal.
Rays mostly 3–4 cells wide, uniseriate rays not common ( Fig. 4F, G View Figure 4 ). Weakly heterocellular. Average multiseriate ray height 583 (212) µm, range 271–1243 µm; 5–8 per mm. Oil/mucilage cells and crystals not observed.
Comparisons with extant woods— Affinities with Magnolia are indicated by this combination of features: diffuse-porous wood with distinct growth rings (1p, 5p), vessels solitary and in radial multiples but without a distinctly patterned arrangement (6a, 7a, 8a, 9a), exclusively scalariform perforation plates with less than 20 bars (13a, 14p, 18a), scalariform intervessel pitting (20p), vessel-ray parenchyma horizontally elongate and with reduced borders (30a), helical thickenings throughout the body of the vessel element (36p, 37p), non-septate fibers (66p), marginal parenchyma (89p), rays not mark- edly heterocellular (108a, 109a), and oil/mucilage cells in rays absent (124a), although many tropical Magnolia species have oil/mucilage cells. In addition to the descriptions provided by Metcalfe and Chalk (1950) and Metcalfe (1987), descriptions of Magnoliaceae woods have been presented in publications devoted to the family: Stark (1954), Canright (1955), Chen et al (1993), as well as in wood anatomical atlases: Kribs (1968), Détienne and Jacquet (1983), Sosef et al. (1998), Carreras and Dechamps (1995), Lemmens et al. (1995), Carreras et al. (2012), Aguilar-Alcantara (2014), Sonsin et al. (2014), Eom (2015) and Itoh et al. (2022). Data from most of these publications are in the InsideWood database and were consulted.
The wood anatomy of Magnolia in the broad modern sense of the genus as well as in its earlier, more narrow circumscription is diverse with variation in these features: perforation plates simple and/or scalariform; intervessel pits opposite and/or scalariform; helical vessel wall thickenings present or absent; rays ranging from narrow (1―2-seriate) to medium width (2―5)-se- riate, and secretory cells (oil/mucilage cells) present or absent. The full range of this diversity was described in detail by Chen et al. (1993) for the Magnoliaceae of China — the center of diversity of the family. Chen et al. (1993) provided a dichotomous key to the genera that were recognized at the time and this fossil from Dietz Hill keys to Michelia L. (1753) with 25 species studied, all now subsumed into Magnolia . Of those species, only Magnolia compressa Maxim. (1872) was returned in the search of InsideWood. Although Chen et al. (1993) concluded that the old paraphyletic genera could mostly be recognized by their wood anatomy, they also showed considerable overlap.
Different authors estimate different numbers of genera and species in the Magnoliaceae ; Figlar and Nooteboom (2004) and Wang et al. (2020) recognize only two gen- era of Magnoliaceae ( Liriodendron L.,1753, Magnolia ), while Xia et al. (2008) and Sima and Lu (2012) divided Magnolia into 16 genera. Estimates of the number of species vary: Stevens (2001 ‒onwards) ca. 225 species of Magnolia ; the Flora of China ca. 300 species; Mabberley (2017) 221 species.
Only 60 extant Magnolia species are represented in InsideWood by descriptions and/or images. Six match the combination of features of the fossil: the Asian Magnolia compressa Maxim. (1872) , M. fordiana (Oliv.) Hu (1924) , M. kwangsiensis Figlar and Noot. (2004) , and M. sieboldii Koch (1853) ; the North American M. grandiflora L. (1759) and M. virginiana L. (1753). No Neotropical Magnolia species match UF 278-84871's features. Not all of the aforementioned species have been included in phylogenetic analyses. Figlar and Nooteboom (2004), as did Wang et al. (2020), placed M. grandiflora and M. virginiana in subgenus Magnolia sect. Magnolia : M. grandiflora is the better match for the fossil as its helical thickenings are distinct. Wang et al. (2020) placed M. fordiana ( S. China to Vietnam) in sect. Manglietia ; M. kwangsiensis ( S. China) in sect. Kmeria (Pierre) Dandy (1927) and the temperate Asian M. sieboldii in sect. Oyama. A review of the Leiden wood slide collection and a new atlas for Chinese woods ( Itoh et al. 2022) showed that M. foveolata (Merr.ex Dandy) Figlar (2000) and Ma. martini H. Lév. (1904) also are close matches for this Dietz Hill wood.
Given the overlap in wood anatomy within Magnolia and there being over 100 species whose wood anatomy is unknown, it is highly likely that M. hansnooteboomii is a close match of more species than the ones discussed above.
Comparisons to fossil woods— Fossil woods of Magnoliaceae date back to the late Cretaceous (Campanian). Among the 18 fossil Magnoliaceae woods in the InsideWood database ( Table 1), two species share the aforementioned features with M. hansnooteboomii : M. nanningensis Huang, Jin and Okolski (2019) from the upper Oligocene, China, and Magnolioxylon parenchymatosum van der Burgh (1973) from the Miocene of western Europe,
Although today Magnoliaceae are no longer native in western North America, fossil woods confirm its for- mer presence, including three representatives from the middle Eocene Clarno Nut Beds of Oregon: M. pageae Wheeler and Manchester (2002) , Magnoliaceoxylon angulata (Scott and Wheeler) Wheeler and Manchester (2002) , and Ma. cutleri Wheeler and Manchester (2002) . All differ from M. hansnooteboomii ( Table 1). Seed casts of Magnolia also occur at the Dietz Hill locality ( Manchester and McIntosh 2007).
The wood that Huard (1967) described as Laurinoxylon intermedium from the Neogene of southwest France is not Lauraceae , but Magnoliaceae . That species had exclusively scalariform perforation plates and scalariform intervessel pitting, which do not occur in the Lauraceae . It seems probable that he used the occurrence of idioblasts to assign the sample to Lauraceae and was unaware that secretory idioblasts also occur in Magnolia wood. Additionally, Huard’s wood does not have paratracheal parenchyma, only widely spaced apotracheal parenchyma bands, yet another feature consistent with magnoliaceous affinities.Accordingly, we hereby propose a new combination, Magnolia intemedia (Huard) comb. nov. Wheeler, Manchester, and Baas.
A comparison with fossil Magnolia s.l. species described in the literature, including M. intermedia , ( Table 1) reveals that UF 278-84871differs in its combination of intervessel pits predominantly scalariform; helical vessel wall thickenings present; oil/mucilage cells absent. This
and Stockey (1990); 2 = Huang et al. (2020; 3 = Huard (1967); 4 = Iamandei et al. (2011); 5 = Iamandei et al. (2020); 6 = Page (1970); 7 = Prakash et al. (1971); 8 = Sakala et al. (2010); 9 = Srivastava and Suzuki (2001); 10 = Suzuki (1976); 11 = Takahashi and Suzuki (2003); 12 = van der Burgh (1973); 13 = Wheeler et al. (1977); 14 = Wheeler and Manchester (2002).
Species Age IVP Bars/PP Si HT OM Reference
Magnoliaceoxylon hokkaidoense K (C-M) S, O 12–47 N N N 11
Magnoliaceoxylon panochensis K ( C)
S, O
up to 10 N N N 6
Magnoliaceoxylon wetmorei e m Eo S, ( O)
8–26 N N Y 13
Liriodendroxylon mutltiporosum m Eo
Liriodendroxylon princetonensis * m Eo
Magnolia pageae m Eo
O
S, O
O
2–10 N N ( Y) 14
8–18 N N N 1
5–20 N N Y 14
Magnoliaceoxylon angulata m Eo S, O Magnoliaceoxylon cutleri m Eo S, ( O)
Magnoliaceoxylon longiradiata m Eo S, ( O)
15–18 N N N 14
8–20 N N Y 14
6–20 N N ( Y) 14
Magnolia hansnooteboomii sp. nov. l Eo
Magnoliaceoxylon palaeogenica e Olig
Liriodendroxylon tulipiferum Olig
Magnolia nanningensis up Olig
Magnolioxylon c f. transilvanicum m Mio
S
S
S, O
O
S, O
S, O
8–16 N Y N This paper
4–15 N N N 9
7–15 N Y Y 10
~10 N N N 7, 8
2–11 N Y Y 2
short Y N N 4
Magnolioxylon kräuselli m Mio ( S), O
10–12 N N N 5
S
15–20 N N Y 3
Magnolioxylon parenchymatosum Mio S 3–12 N N N 12
justified its recognition as a new species.
| SP |
Instituto de Botânica |
| UF |
Florida Museum of Natural History- Zoology, Paleontology and Paleobotany |
| L |
Nationaal Herbarium Nederland, Leiden University branch |
| S |
Department of Botany, Swedish Museum of Natural History |
| A |
Harvard University - Arnold Arboretum |
| OM |
Otago Museum |
| K |
Royal Botanic Gardens |
| O |
Botanical Museum - University of Oslo |
| N |
Nanjing University |
| C |
University of Copenhagen |
| Y |
Yale University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |