
Mesalina arnoldi, Sindaco & Simó-Riudalbas & Sacchi & Carranza, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4429.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:2FE8E74C-6ABF-4EA9-A7B6-EA4E2525ABC4 |
|
DOI |
https://doi.org/10.5281/zenodo.5951784 |
|
persistent identifier |
https://treatment.plazi.org/id/038B87D9-5D06-FFA5-FF57-783FD80AF84C |
|
treatment provided by |
Plazi |
|
scientific name |
Mesalina arnoldi |
| status |
sp. nov. |
Mesalina arnoldi sp. nov.
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 , 6 View FIGURE 6 ; Tables 1–5, Appendices I and III)
Mesalina sp. A Arnold 1986a: 427, Schätti & Gasperetti 1994: 371; Mesalina guttulata Sindaco & Jeremcenko 2008: 262 (part.).
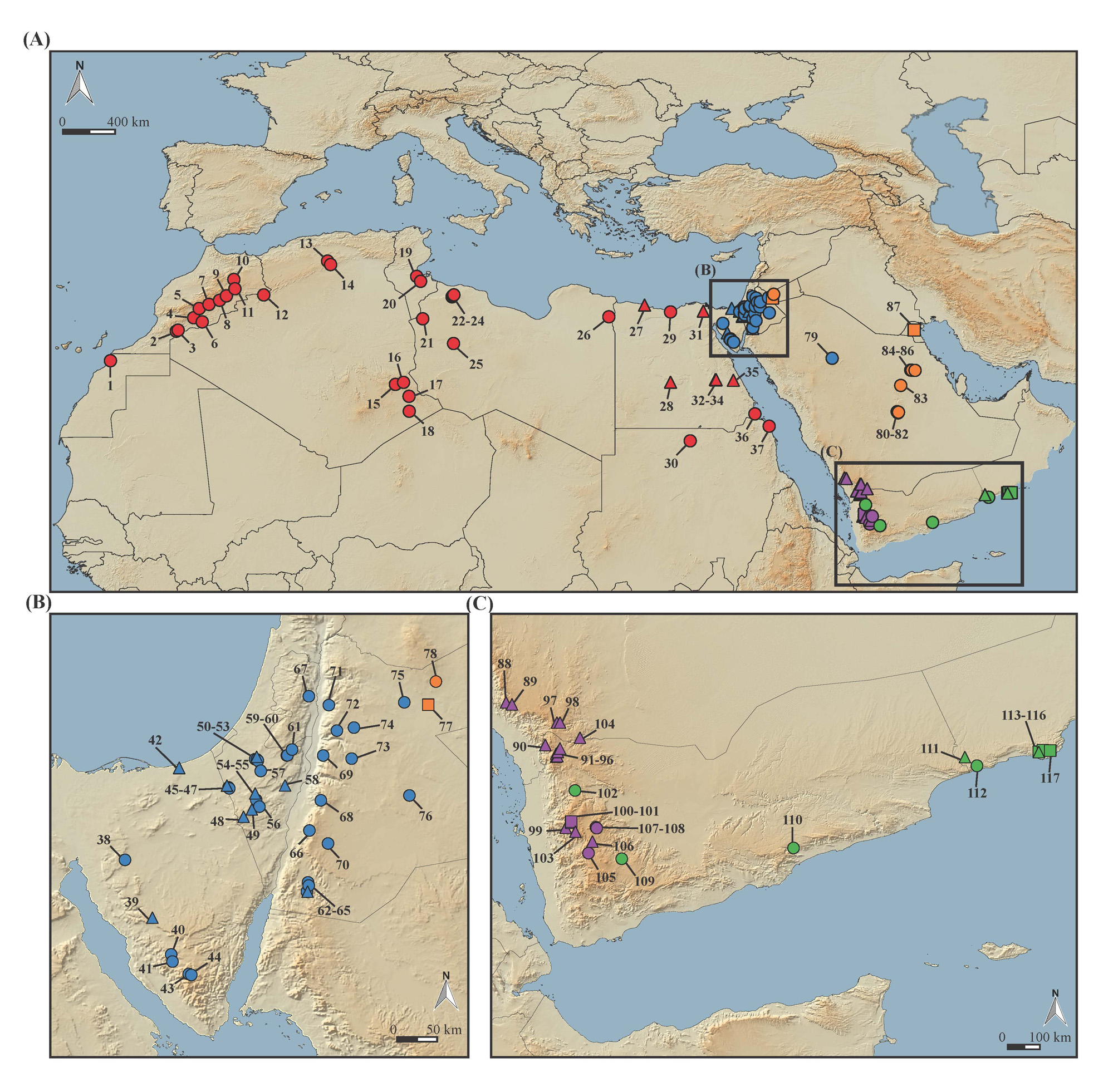
Holotype. Adult female MCCI-R890 , Yemen, Amran Governatorate, plateau between Zakatin village ( Hababah ) to Kawkaban ( Haraz Mt. ) (about 15.51°N, 43.86°E WGS84), 2,600-2,800 m a.s.l., R. Sindaco and C. Sindaco leg., 7 February 1998. GoogleMaps
Paratype. Adult male MZUF-28670, Yemen, Al Mahwit Governatorate, Kawkaban (about 15.50°N, 43.90°E WGS84), M. Poggesi, M. Borri, M. Manetti and M. Sammicheli leg., 31 January 1984. GoogleMaps
Other specimens examined. Forty-four specimens in the collections of the Natural History Museum in London and in the Museum “La Specola” in Florence (see Appendix I) plus four specimens used only for genetic analyses (no vouchers available, juvenile or damaged specimens); all listed in Appendix I.
Etymology. The species epithet “ arnoldi ” is a genitive Latin noun to honor the British herpetologist Dr E. Nicholas Arnold for his life-long dedication and contribution to Arabian herpetology, including the recognition of this taxon as a distinct species that he provisionally referred to as Mesalina sp. A in Arnold (1986a).
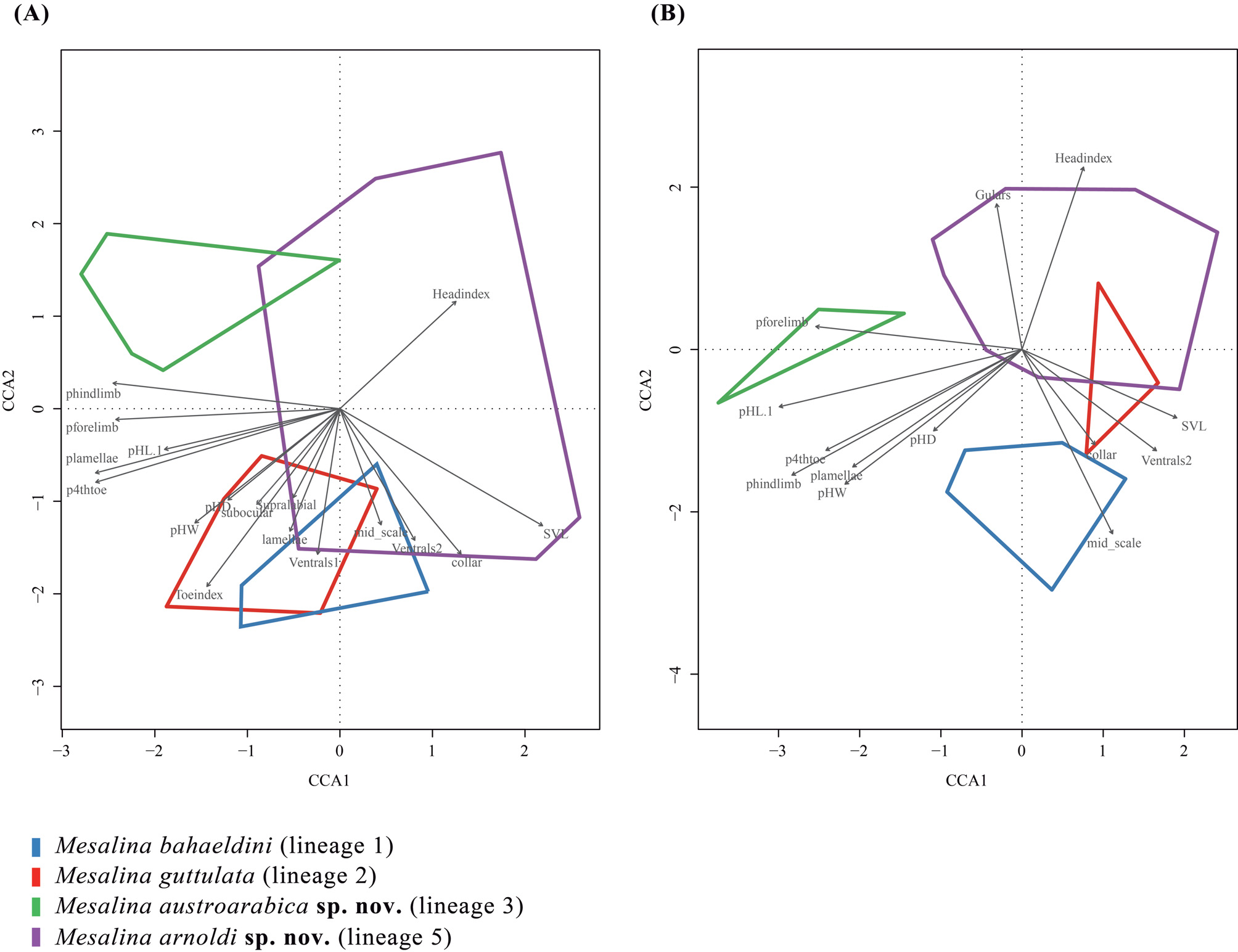
Diagnosis. A relatively large-sized Mesalina characterized by the following combination of morphological characters: (1) well-developed occipital scale in contact with the interparietal (with rare exceptions) ( Fig. 6E View FIGURE 6 ); (2) lower eyelid with a window made of up two large scales (in 57% of examined specimens) or fragmented into smaller scales (43%) ( Fig. 6D View FIGURE 6 ), often without black edges (67%); (3) curved collar ( Fig. 6F View FIGURE 6 ); (4) four upper labials in front of the subocular in 89% of the samples and five in 11% of the samples ( Fig. 6D View FIGURE 6 ); (5) ventral plates in 10 (very rarely 8) straight longitudinal rows, the outermost much smaller ( Fig. 6B View FIGURE 6 ); (6) scales on the upper surface of the tibia keeled ( Fig. 6A View FIGURE 6 ); (7) lamellae under 4th toe, 19–26 (median = 22); (8) dorsal pattern usually very marked, background color brown-greyish, with many complete ocelli (i.e. a white spot completely surrounded by a black ring) or near so, ordered in irregular longitudinal and transverse rows. Dorsolateral and light stripes are usually evident, often interrupted; some specimens are clearly striped, while in others these lines are inconspicuous, only rarely absent ( Fig. 6A View FIGURE 6 )
There are no obvious morphological characters separating M. arnoldi sp. nov. from M. guttulata , M. bahaeldini and M. austroarabica sp. nov. The statistical analyses (see Results above) show significant differences, with M. arnoldi sp. nov. having more gulars (males), more dorsals at midbody (males), more femoral pores (males and females) than M. guttulata . Moreover, M. arnoldi sp. nov. has smaller %HD (males), smaller %4th toe length (males), smaller toe-index (males), lesser value of Lamellae percSVL (males). Mesalina arnoldi sp. nov. shows significant differences from M. bahaeldini in having more gulars (males and females), more femoral pores (males) and more supralabials (females). Differences between M. arnoldi sp. nov. and M. austroarabica sp. nov. are discussed in the description of the latter species (see above).
Genetic and phylogenetic remarks. This species had not been included in any previous phylogenetic analyses, not even the comprehensive study by Kapli et al. (2015). The phylogenetic analyses performed in this study ( Fig. 2 View FIGURE 2 ; Table 1) support the hypothesis that M. arnoldi sp. nov. is an independent species. The level of genetic differentiation ( p -distance) between the new species and the other members of the Mesalina guttulata species complex ranges between 5.2–6.6% in the 12S, 6.1–7.1% in the 16S and 11.9–15.7% in the cytb genes ( Table 1). A network analysis of the nuclear gene MC1R indicates that, despite the large number of samples of the M. guttulata species complex included in the analysis ( 36 specimens; 72 alleles), all four haplotypes (10 alleles) of M. arnoldi sp. nov. are private ( Fig. 3 View FIGURE 3 ; Appendix I).
Description of the holotype. An adult female with partly regenerated tail. Measurements, meristic characters and indexes: SVL = 53.0 mm, HL-1 = 11.7 mm (22% of SVL), HL-2 = 5.0 mm (9% of SVL), HL-3 = 4.3 mm (8% of SVL), Head width = 7.3 mm (14% of SVL), Head depth = 4.9 mm (9% of SVL), pileus = 10.4 mm (20% of SVL), Forelimb length = 16.3 mm (31% of SVL), Hindlimb length = 26.6 mm (50% of SVL), 4th toe length = 8.6 mm (16% of SVL), Tail length = 62.0 mm (partly regenerated), supralabials 9/9, subocular = 5/5, gulars = 23, enlarged plates in collar = 9, midbody scales = 48, longitudinal rows of ventrals = 8+2 (smaller), transversal rows of ventrals = 36, femoral pores = 14+14, lamellae under the 4th toe = 21. Head index = 160, Toe index = 32, Lamellae percSVL = 0.77.
Coloration in alcohol: numerous ocelli, each one formed by several whitish scales forming a dot and surrounded by an almost complete ring of black colored scales (ocelli are reduced to black dots on the neck). These ocelli form 4-6 rather regular longitudinal series (the paravertebral and lateral ones more marked) and about 13 very irregular transverse series, between fore- and hindlimbs; black and white dots are present on the tail and hindlimbs. The pileus is grey without specks (only the outer margin of the parietals is bordered with black). On the sides of the head a continuous dark stripe is present from near the upper border of the ear opening, across the eye, to the loreal scale. Another well-defined dark stripe lies between the mid- ear opening and the subocular scale. Flanks with two series of ocelli, without evident stripes. The ventral side is creamy-white, immaculate, with the exception of infralabial scales, irregularly sprinkled with gray, as well as outer ventrals and the anterior margin of thighs.
Variation. In specimens MZUF-28132 the occipital scale is almost absent. The two lateral rows of ventrals are usually much smaller than the inner ventrals, sometimes subequal in size, and absent in specimen MZUF-28132. The dorsal pattern is very variable; specimens with the pattern similar to the holotype are frequent, but in several specimens the white dots of outer dorsal ocelli tend to form a whitish, more or less interrupted, supraciliar stripe along the sides of the back. In several specimens, instead of small ocelli, there are dark blotches on the back, parallel to the light supraciliar stripes, forming a distinct striped pattern (BM1938.8.1.27, BM1977.423). In specimens BM1977.425 and MZUF-28673 there are four uninterrupted white stripes: two supraciliar stripes and two subocular stripes along the sides of the body.
Coloration in life. Ground color brownish with more or less intense shades of gray. Ocelli whitish surrounded by dark brown incomplete rings ( Fig. 6G View FIGURE 6 ).
Distribution and habitat. Specimens referable to Mesalina arnoldi sp. nov. are widespread in the highlands of southwestern Saudi Arabia and Western Yemen. The holotype was collected in a stony plateau with basaltic rocks and scarce vegetation, at an altitude of 2,600-2,800 m a.s.l. The paratype was collected in the same area, between 1,950 and 2,300 m a.s.l. According to Schätti & Gasperetti (1994) this species is found as low as 1,300 m a.s.l.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.