
Diorhabda meridionalis Berti & Rapilly, 1973
|
publication ID |
https://doi.org/10.11646/zootaxa.2101.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/038C2C5B-AC1A-FFC2-3B91-FA1CF7ADD66A |
|
treatment provided by |
Felipe |
|
scientific name |
Diorhabda meridionalis Berti & Rapilly, 1973 |
| status |
stat. nov. |
Diorhabda meridionalis Berti & Rapilly, 1973 NEW STATUS southern tamarisk beetle
( Figs. 9 View FIGURES 1–9 , 18 View FIGURES 14–18 , 23 View FIGURES 19–23 , 28 View FIGURES 24–28 , 33 View FIGURES 29–33 , 38 View FIGURES 34–38 , 43 View FIGURES 39–43 )
Diorhabda carinulata meridionalis Berti & Rapilly, 1973:881 ( Type locality: Minab, Iran); Warchalowski, 2003:328 (catalog of Mediterranean Region, Iran).
Diorhabda elongata: Riley et al., 2003 :69,189 (part, catalog of North America [not yet introduced]).
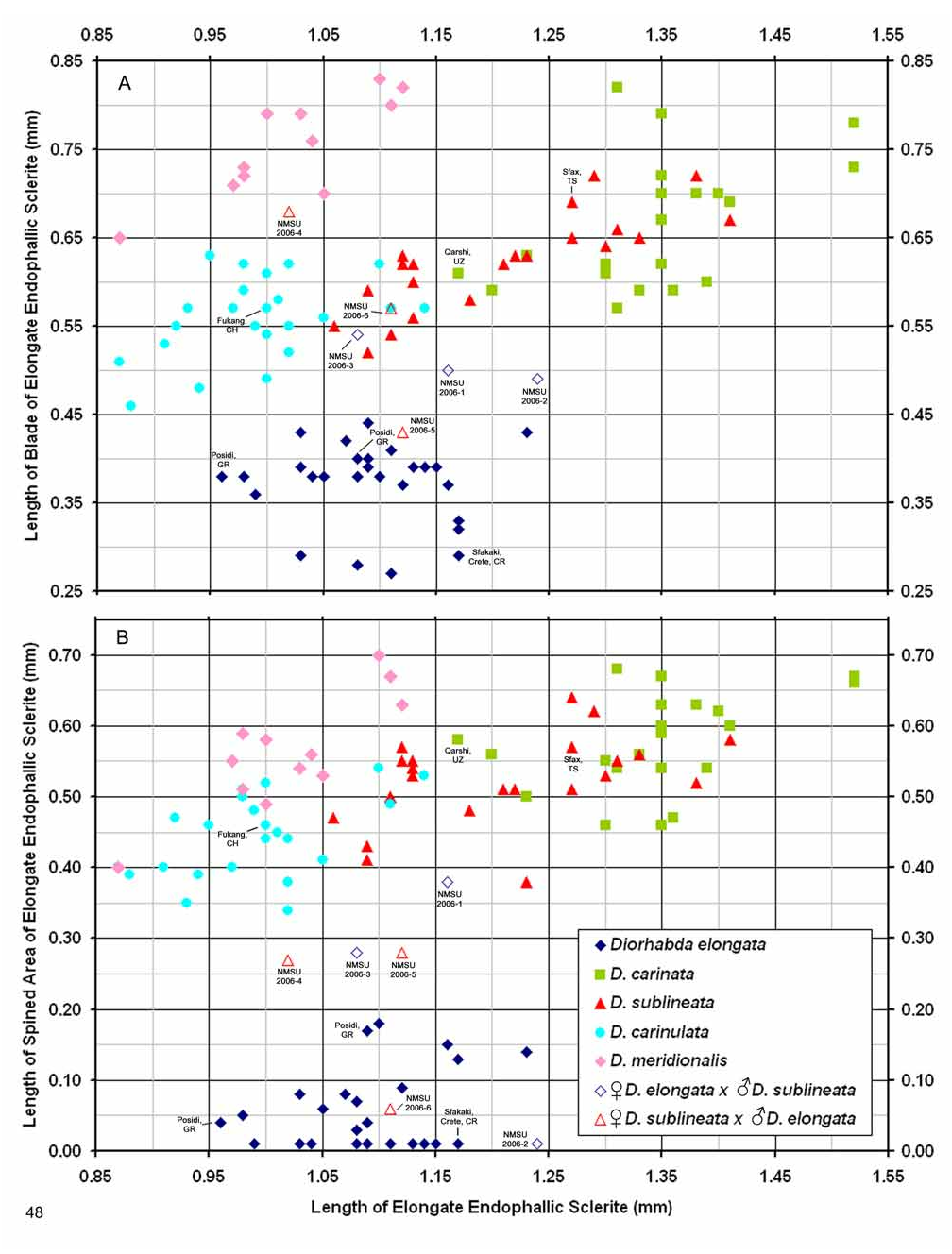
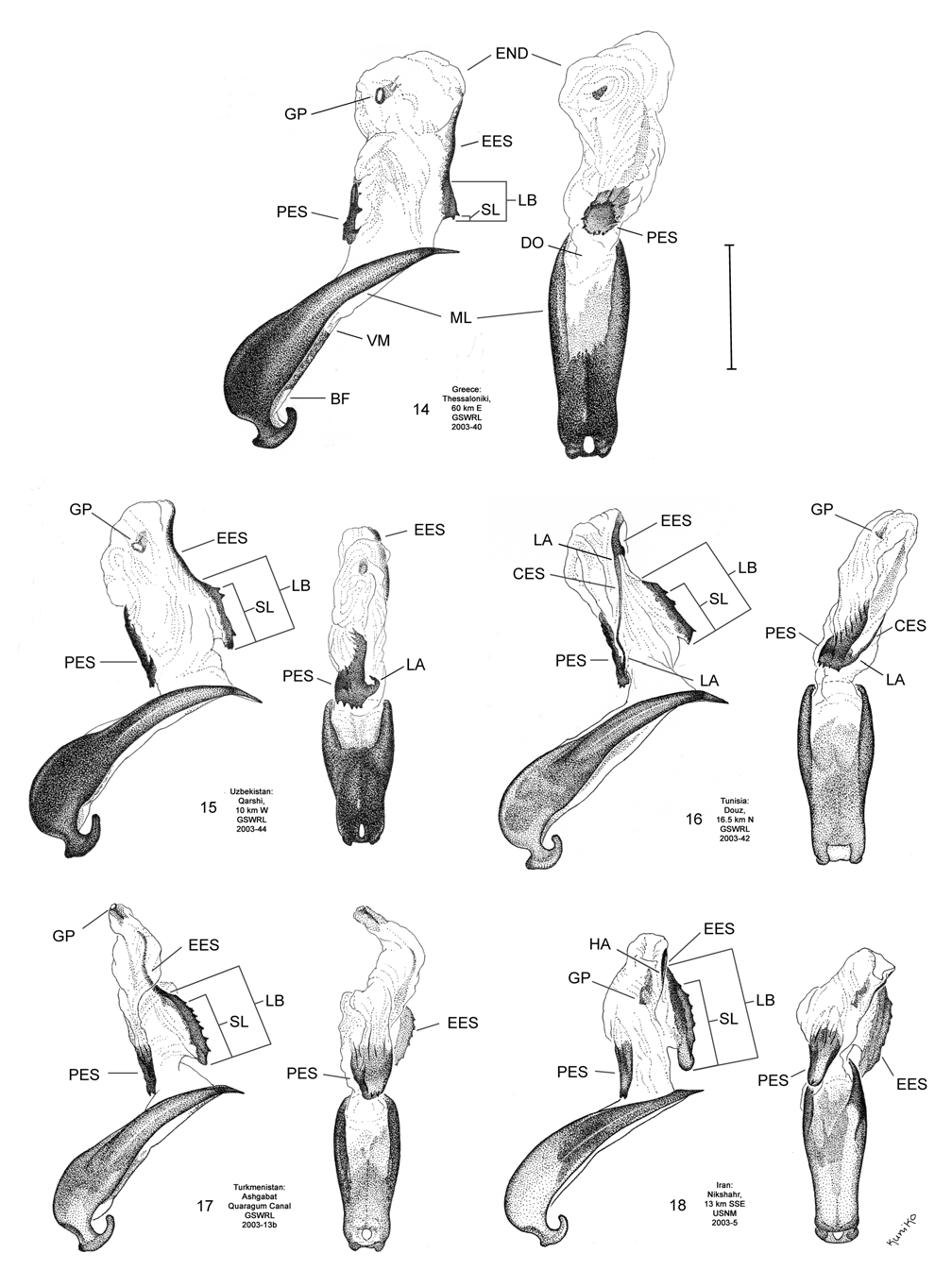
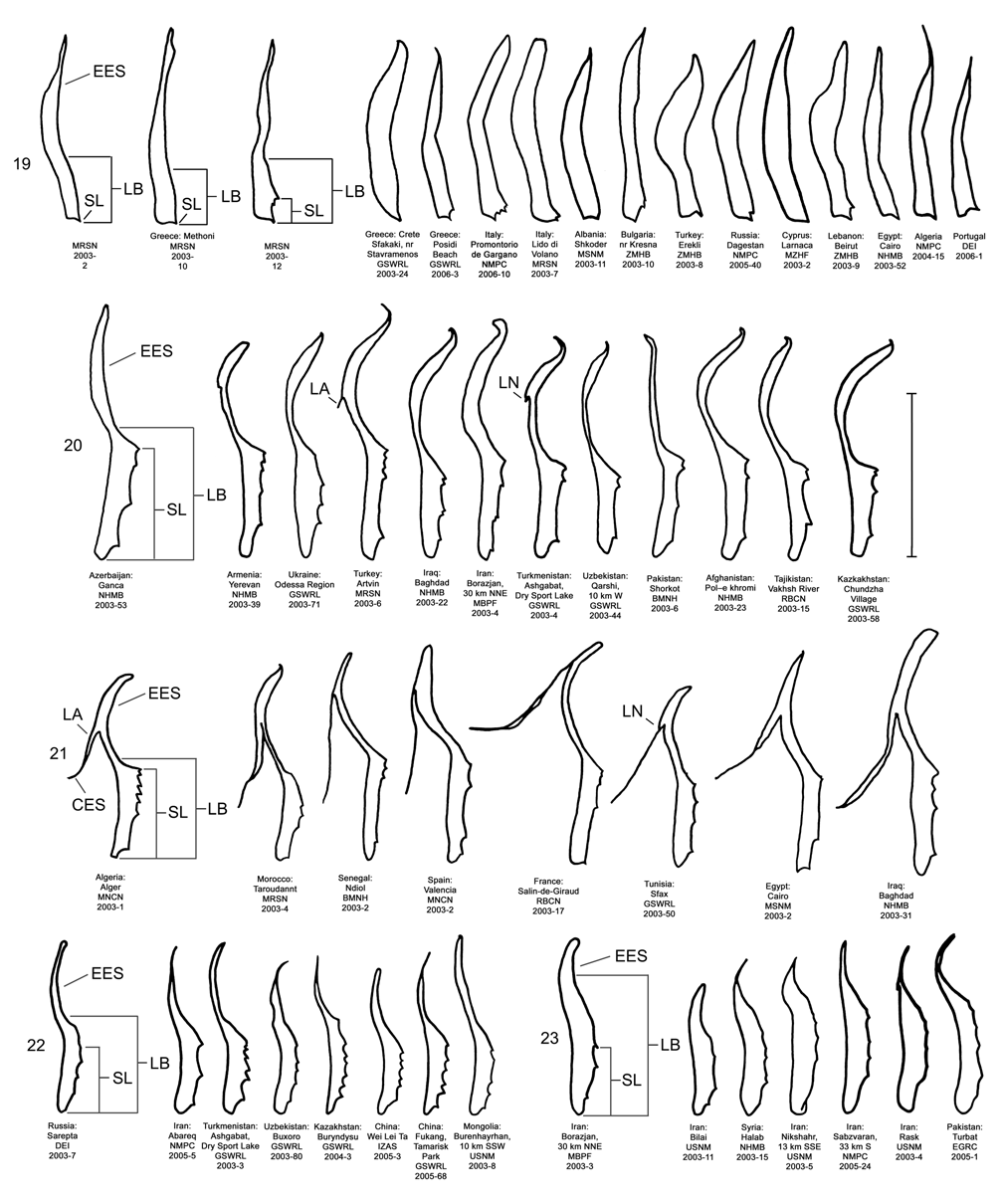
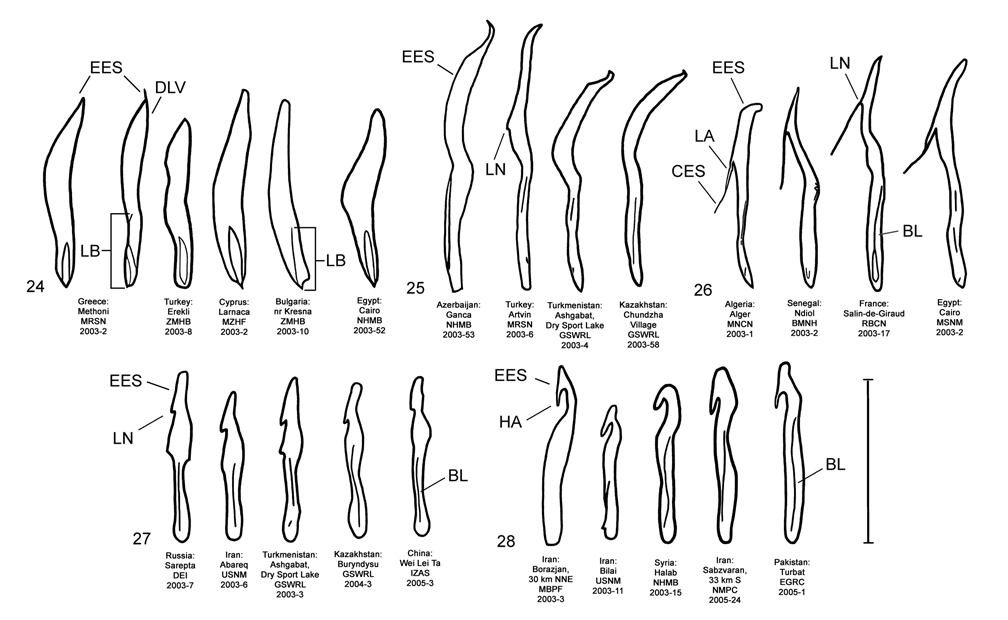
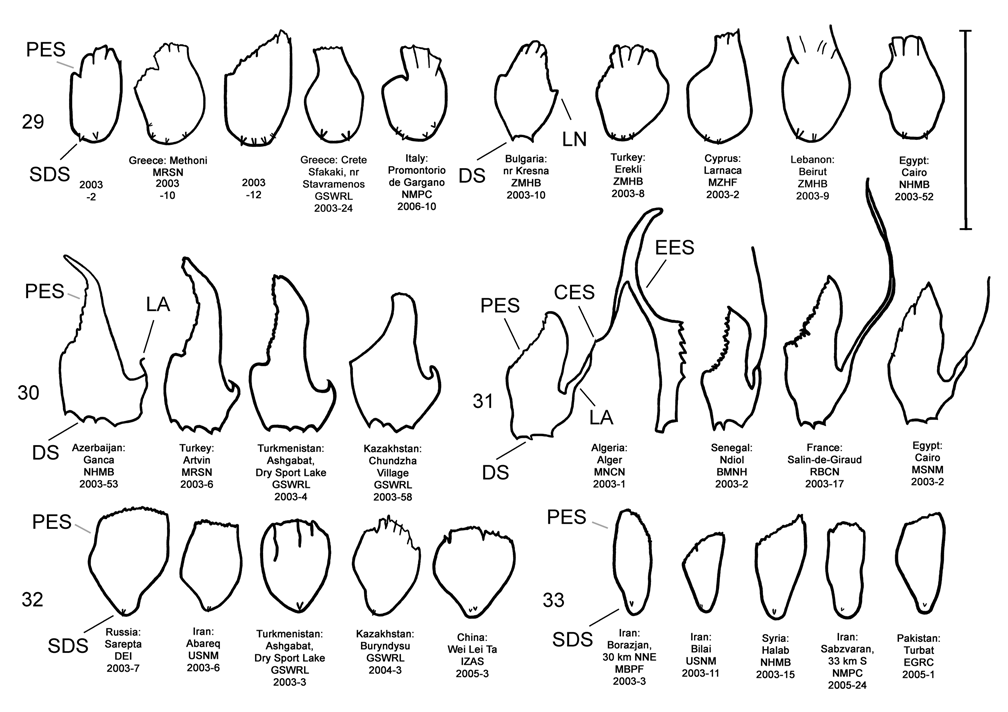
Male. Genitalia. Diorhabda meridionalis can be distinguished from all other members of the D. elongata group by two unique characteristics of the elongate endophallic sclerite (EES): (1) the presence of a strongly hooked apex ( Figs. 18 View FIGURES 14–18 , 28 View FIGURES 24–28 ), and (2) the length of the blade of the EES being greater than or equal to 0.67 times the length of the EES ( Table 3; Figs. 18 View FIGURES 14–18 , 23 View FIGURES 19–23 , 48 View FIGURE 48 ). In all other members of the D. elongata group, the blade is less than or equal to 0.66 times the length of the EES ( Figs. 14–17 View FIGURES 14–18 , 19–22 View FIGURES 19–23 ) and the apex of the EES is not hooked ( Figs. 24–27 View FIGURES 24–28 ). The palmate endophallic sclerite (PES) of D. meridionalis is also unique among the D. elongata group in that its distal margin is narrowly rounded and generally smooth, with only one or two small subdistal spines, and it has a length to width ratio of 0.35–0.63 ( Figs. 18 View FIGURES 14–18 , 33 View FIGURES 29–33 ; Table 3). In contrast, the distal margin of the PES is truncate or more broadly rounded with larger distal or subdistal spines in D. elongata , D. carinata , and D. sublineata ( Figs. 14–16 View FIGURES 14–18 , 29–31 View FIGURES 29–33 ), and the PES is acutely rounded with a length to width ratio of 0.61–1.02 in D. carinulata ( Figs. 17 View FIGURES 14–18 , 32 View FIGURES 29–33 ).
Measurements. See Tables 2 and 3.
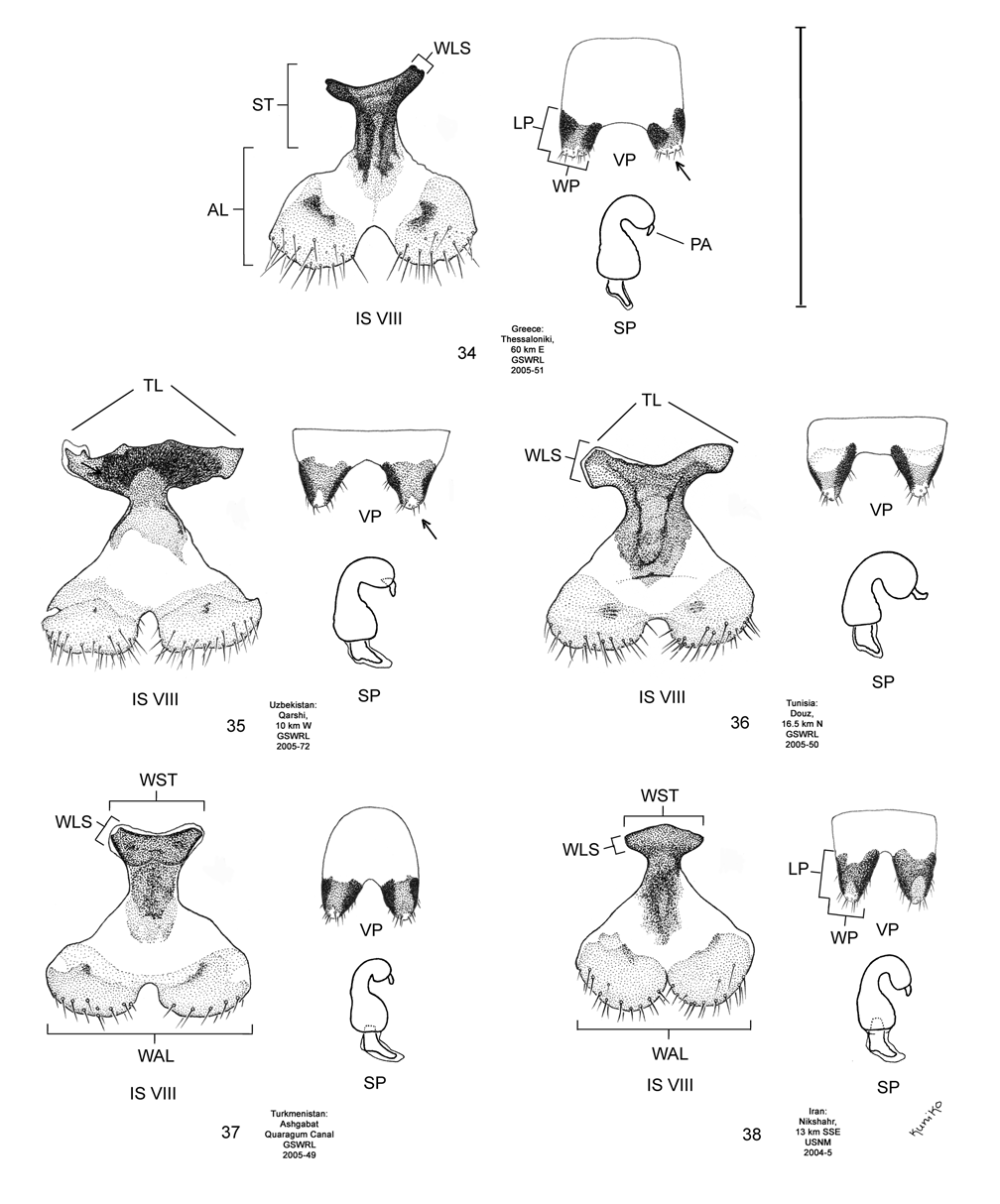
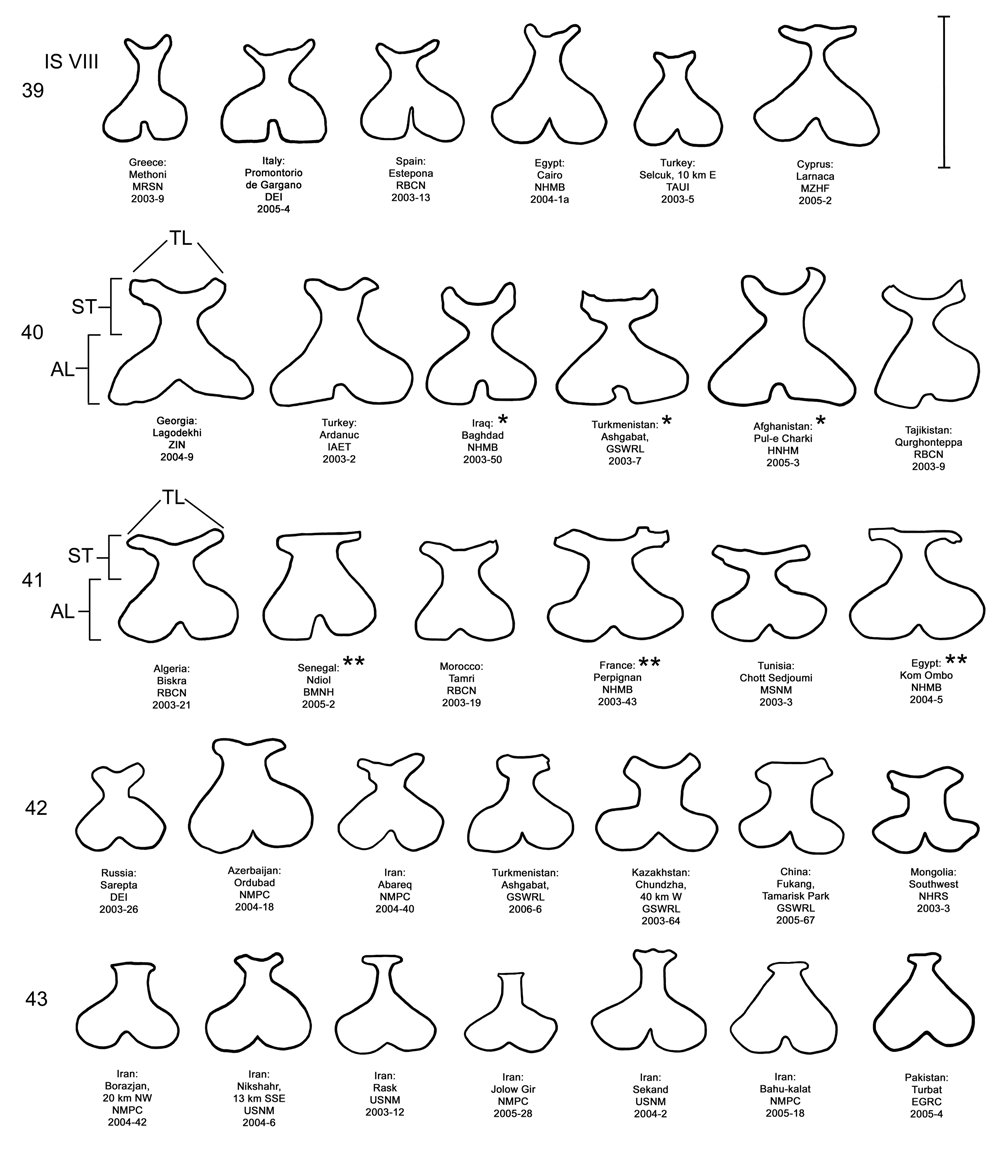
Female. Genitalia. Female D. meridionalis can be distinguished from all other members of the D. elongata group by the following combination of characters in the vaginal palpi (VP) and internal sternite VIII (IS VIII): (1) the vaginal palpi are broadly rounded and as long as wide or longer with a width to length ratio (LP/WP) of 1.0–1.31 ( Fig. 38 View FIGURES 34–38 ; Table 4), and (2) the width of the stalk (WST) of IS VIII is 0.22–0.33 mm and this width is 0.33–0.48 times the width of the apical lobe (WAL) ( Figs. 38 View FIGURES 34–38 , 43 View FIGURES 39–43 ; Table 4). In contrast, the vaginal palpi are wider than long, with a length to width ratio of 0.46–0.94 in D. elongata , D. carinata and D. sublineata ( Figs. 34–36 View FIGURES 34–38 ). In addition, the vaginal palpi are triangulate in D. carinata and D. sublineata ( Figs. 35–36 View FIGURES 34–38 ). In D. carinulata , the width of the stalk of IS VIII is 0.36–0.57 mm and this width is from 0.49–0.77 times the width of the apical lobe ( Fig. 37 View FIGURES 34–38 , 42 View FIGURES 39–43 ). In addition, the width of the widest lobe of the stalk of IS VIII is from 0.04–0.09 mm in D. meridionalis ( Fig. 38 View FIGURES 34–38 , 43 View FIGURES 39–43 ); whereas, in D. carinulata the width of the widest lobe of the stalk is usually greater than 0.10 mm ( Figs. 37 View FIGURES 34–38 , 42 View FIGURES 39–43 ; Table 4).
Measurements. See Tables 2 and 4.
Coloration. Coloration of dead D. meridionalis ( Fig. 9 View FIGURES 1–9 ) is similar to that discussed above for D. sublineata and D. carinulata ( Figs. 5, 7 View FIGURES 1–9 ). We have not had the opportunity to examine live material of D. meridionalis .
Type material. Berti and Rapilly (1973) deposited the holotype male and allotype female of D. meridionalis , both from Minab, Iran, and four paratypes ( 3♂♂, 1♀) from Minab, Borazjan and Shush (= Susa), Iran at MNHN. As discussed above (see D. elongata - Type Material), we were unable to obtain type materials from MNHN. We studied the original description by Berti and Rapilly (1973), including their illustrations of the endophallus of the male holotype. We also examined a male specimen from 30 km NNE of Borazjan, a paratype locality, with an identification label as “ D. carinulata meridionalis Berti & Rapilly det. Berti X-1996 ” ( MBPF 2003-03 ) and additional material near type and paratype localities .
Material examined. 22♂♂ dissected (diss.), 19♀♀ diss., 13♂♂, 18♀♀, 147 unsexed specimens. IRAN: 2♂♂ diss., 3♀♀ diss., 11♂♂, 9♀♀, Bahu–Kalat [ 25.73330°N, 61.53330°E; 68 km south Rask, Sarbaz River; T. dioica and T. aphylla present ( Hoberlandt 1981)], 3–4-IV-1973, Exp. Nat. Mus. Praha Loc. No. 147, NMPC [2004-48, 2005-16, 17, 18, 19]; 3♂♂ diss., Bilai [ 26.48333°N, 57.15000°E; coastal savanna; Tamarix sp. not recorded ( Hoberlandt 1981)], 23–24-V-1973, Exp. Nat. Mus. Praha Loc. No. 209, Diorhabda elongata det. I.K. Lopatin 1978 [ 1♂ diss., USNM], NMPC [2004-57, 2005-22], USNM [2003-11]; 1♂ diss., 2♀♀ diss., 1♀, Borazjan, 20 km northwest [ 29.38333°N, 51.03333°E; Shabankareh, clay semi-desert; Tamarix sp. not recorded ( Hoberlandt 1983)], 18-IV-1977, Exp. Nat. Mus. Praha Loc. No. 297, NMPC [2004-05, 42, 43]; 3♂♂ diss., 1♂, 3♀♀, Borazjan, 30 km north northeast [ 29.53333°N, 51.40000°E; 10 km N Dalaki, Hableh Rud River; Tamarix sp. present ( Hoberlandt 1983)], 18–19-IV-1977, Exp. Nat. Mus. Praha Loc. No. 298, D. carinulata meridionalis Berti & Rapilly det. Berti X-1996 [ 1♂ diss., MBPF], NMPC [2004-12, 2005-20], MBPF [2003-03]; 2♀♀ diss., Jeiugir ( 33° 27' N, 49° 01' E) [sic; Jolow Gir; 32.96667°N, 47,81611°E; coordinates corrected from relation to Sarab–e Jahangir], 20 km southeast Sarab–e Jahanagir, Lorestán, 8–10- X-1998, Chvojka, D. elongata det. A. Warchalowski, NMPC [2005-28, 2006-01]; 1♂ diss., 2♀♀ diss., 1♀, Nikshahr, 13 km south southeast [ 26.13333°N, 60.18333°E; Rudkhaneh Nikshahr river; T. dioica present ( Hoberlandt 1981)], 8–9-IV-1973, Exp. Nat. Mus. Praha Loc. No. 152, USNM [2003-05, 2004-05, 06], NMPC; 1♂ diss., 1♂, 3♀♀, Omidiyeh, 34 km southeast [ 30.55000°N, 49.91667°E; Rud-e Zohreh river; Tamarix sp. present ( Hoberlandt 1983)], 16–17-IV-1977, Exp. Nat. Mus. Praha Loc. No. 292, NMPC [2004- 44]; 8♂♂ diss., 5♀♀ diss., 1♂, 2♀♀, 142 specimens [ NMPC], Rask [ 26.23528°N, 61.40111°E; 3 km N Rask; T. dioica present ( Hoberlandt 1981)], Sarbaz River valley, 3–4-IV-1973, Exp. Nat. Mus. Praha Loc. No. 146, D. carinulata meridionalis det. Lopatin 1984 [ 1♂, 1♀, NMPC], D. elongata det. I.K. Lopatin [ 2♀♀, USNM], USNM [2003-04, 12, 13, 2004-03; 2008-02, 03, 04, 05, 06], NMPC [2004-58, 2005-21, 2006-8–9]; 1♂ diss., 2♀♀ diss., 1♀, Sabzevaran [Sabzvaran], 33 km south [ 28.40000°N, 57.83333°E; ruderal plants near plantation; Tamarix sp. present ( Hoberlandt 1983)], 17-V-1977, Exp. Nat. Mus. Praha Loc. No. 335, NMPC [2004-27, 2005-23, 24]; 1♀ diss., Sekand [ 26.71667°N, 61.53330°E; oasis of saline marshes; Tamarix sp. not recorded, Frankenia pulverulenta present ( Hoberlandt 1981)], 27 km east northeast Sarbaz, 31-III to 1-IV- 1973, Exp. Nat. Mus. Praha Loc. No. 144, USNM [2004-02]; 1♀ diss., Shushtar [ 32.05°N, 48.85°E; river Karun; Tamarix sp. present ( Hoberlandt 1983)], 13-IV-1977, Exped. Nat. Mus. Praha Loc. No. 287, NMPC [2005-35]; PAKISTAN: 1♂ diss., 1♀ diss., Turbat [ 26.02780°N, 63.05060°E], west Balochistan, 8–19-IV- 1993, S. Becvar, EGRC [2005-01, 04]; SYRIA: 1♂ diss., Alep [po] [ Halab; 36.20278°N, 37.15861°E], Syrie, NHMB [2003-15].
Distribution. General. Diorhabda meridionalis is primarily known from extreme southern Iran and along the southern portion of Iran’s border with Iraq. Its native range is from Syria to western and southern Iran and southern Pakistan (Map 5). Additional surveys should be made for D. meridionalis in eastern Iraq and along the western coast of the Persian Gulf in Kuwait, Saudi Arabia, United Arab Emirates, Oman, and east to northern India (following the distribution of a suspected host T. dioica Roxburgh ex Roth , see below; Map5).
Confirmed Records. We confirm the presence of D. meridionalis in Iran (Berti and Rapilly 1973) (see above Material examined) (Maps 5–6). Although D. meridionalis , D. carinulata and D. carinata occur on tamarisk throughout Iran (Map 6), Barkhodari et al. (1981) recorded no Diorhabda in their detailed survey of the Tamarix entomofauna in Iran. This is evidence of a possibly sporadic nature of tamarisk beetle populations in Iran.
New Records. Pakistan and Syria are new distribution records for D. meridionalis (Map 5).
Unconfirmed Records. We find no country distribution records that remain unconfirmed. We have not examined specimens from the following locations of D. meridionalis in Iran reported by Berti and Rapilly (1973) (Map 5):
IRAN: 1♂, 1♀, Borazjan [ 29.26222°N, 51.20528°E], 24-V-1969, R. Naviaux and M. Rapilly, on Tamarix sp. , PARATYPE, MNHN GoogleMaps ; 2♂, 1♀, Minab [ 27.14667°N, 57.07361°E], 25-IV-1971, Naviaux and M. Rapilly, on Tamarix sp. , HOLOTYPE, MNHN GoogleMaps ; 1♂, Susa [Shush; 32.19417°N, 48.24361°E], 1899, Escalera, PARATYPE, MNHN.
Discussion. Taxonomy. Diorhabda carinulata meridionalis was described from Minab, Iran with paratype localities in Shush and Borazjan, all along the southwestern border of Iran (Berti and Rapilly 1973). We examined one male specimen from 30 km NNE of Borazjan, a paratype locality, with an identification label as “ D. carinulata meridionalis Berti & Rapilly det. Berti X-1996 ” (MBPF 2003-03) with endophalli matching that of the description by Berti and Rapilly (1973). Other males with endophalli matching that of D. c. meridionalis were examined from Bilai, 74 km NNW from the type locality of Minab, from 20 km NNW of Borazjan, and from six other locations along the southwestern Iranian border (Map 5). Three key characters of the endophallus of D. c. meridionalis that distinguish it from other members of the D. elongata group can be seen in the illustrations of the endophallus of the holotype ( Fig. 19d–e View FIGURES 19–23 of Berti and Rapilly 1973) and our figures ( Figs. 18 View FIGURES 14–18 , 23 View FIGURES 19–23 , 28 View FIGURES 24–28 , 33 View FIGURES 29–33 ): (1) narrowly rounded distal margin of the palmate endophallic sclerite; (2) elongate endophallic sclerite (EES) with blade extending for more than 2/3 the length of the sclerite; and (3) the hooked apex of the EES. We are certain that specimens we studied with endophalli matching that of D. c. meridionalis are conspecific. We find additional characters in the endophallic sclerites ( Figs. 18 View FIGURES 14–18 , 23 View FIGURES 19–23 , 28 View FIGURES 24–28 , 33 View FIGURES 29–33 ) and female genitalia (vaginal palpi and internal sternite VIII; Figs. 38 View FIGURES 34–38 , 43 View FIGURES 39–43 ) of D. c. meridionalis throughout its range (Map 5) that distinguish it from other members of the D. elongata group (see Male- Genitalia and Female- Genitalia above; Figs. 14–17 View FIGURES 14–18 , 19–22 View FIGURES 19–23 , 24–27 View FIGURES 24–28 , 29–32 View FIGURES 29–33 , 34–37 View FIGURES 34–38 , 39–42 View FIGURES 39–43 ). If D. c. meridionalis were a subspecies of D. carinulata , we would expect intermediate forms to occur in areas where their distributions approach one another in southern Iran. Diagnostic characters of the endophalli and female vaginal palpi of D. c. meridionalis are distinct from that of D. carinulata , even in the populations at Sabzvaran, Iran, that approach within 102 km of D. carinulata at Abaraq, Iran (see Biogeography below; Maps 1 and 5, Table 8). The lack of intermediate forms between the two taxa is strong evidence of their reproductive isolation. The distinctive genitalic characters of D. c. meridionalis are also maintained in the same areas where D. carinata occurs and near the abutting range boundary of D. c. meridionalis with D. elongata , and this is strong evidence for reproductive isolation between these species. Although D. c. meridionalis is allopatric with D. sublineata , their status as reproductively isolated is supported by the number of diagnostic genitalic characters separating D. c. meridionalis from D. sublineata (8 characters) being greater than the number of characters separating the moderately sympatric D. carinata from D. carinulata (5 characters) ( Table 5). Therefore, we elevate D. c. meridionalis to species status as D. meridionalis Berti and Rapilly NEW STATUS.
We were unable to fully evert the tip of the endophallus containing the gonopore in D. meridionalis without damaging the specimens ( Fig. 18 View FIGURES 14–18 ), and a more fully everted pointed tip can be seen in the illustration of the holotype endophallus in Fig. 19d–e View FIGURES 19–23 of Berti and Rapilly (1973). We have examined specimens of D. meridionalis that were misidentified by taxonomists as D. elongata from Iran (see Material Examined).
Common Name. The vernacular name “southern tamarisk beetle” refers to its Latin name “ meridionalis ” and its southern range in relation to the closely related and parapatric D. carinulata ( Fig. 51B View FIGURE 51 ).
Biology. Host Plants. Berti and Rapilly (1973) report Tamarix sp. as a host plant for D. meridionalis (as D. c. meridionalis ) in Iran ( Table 1). Records of Tamarix identified to species and other plants present were made in conjunction with collections of D. meridionalis in southeastern Iran by the second Czechoslovak- Iranian Entomological Expedition in 1973 ( Hoberlandt 1981; see above Material Examined). We examined D. meridionalis from one of two expedition collection sites with both T. dioica and T. aphylla (Bahu–kalat), two of five sites with T. dioica and no T. aphylla (Rask and 13 km SSE Nikshahr), and none of eight sites with T. aphylla and no T. dioica . Based on these records, T. dioica , a close relative of T. aphylla , is a probable host plant in southeastern Iran. We found no reports of T. dioica in western Iran and Syria ( Schiman-Czeika 1964, Baum 1978, Zieliṅski 1994), and other Tamarix spp. probably serve as hosts of D. meridionalis in these areas (Maps 5 and 6). Tamarix aphylla is common in southern Iran and southern Pakistan ( Browicz 1991) where it might serve as a host for D. meridionalis .
At the time of collection of D. meridionalis in an oasis of small saline marshes at Sekand, Iran, Hoberlandt (1981) recorded as present the annual herb Frankenia pulverulenta Linnaeus (of the order Tamaricales with Tamarix ) but not Tamarix . Frankenia pulverulenta is indigenous to the southern Palearctic region and southern Africa ( Jäger 1992), and it sometimes grows in association with Tamarix in north Africa (Kassas and Imam 1954), southern Europe ( Cano et al. 2004), and probably also in Iran. It is unlikely that this herbaceous Frankenia would serve as a host of D. meridionalis , but in future surveys for D. meridionalis in the region, F. pulverulenta should be surveyed in order to rule it out as a host.
Diorhabda meridionalis and D. carinata have been collected together in series from five locations in western Iran and Syria, and they probably share some of the same host Tamarix spp. in a manner similar to D. carinulata and D. carinata .
Ecology and Phenology. Collection dates in Iran and Pakistan for D. meridionalis from examined material and collections records of Berti and Rapilly (1973) are from 31 March through 10 October.
Natural Enemies. We found no reports of natural enemies of D. meridionalis .
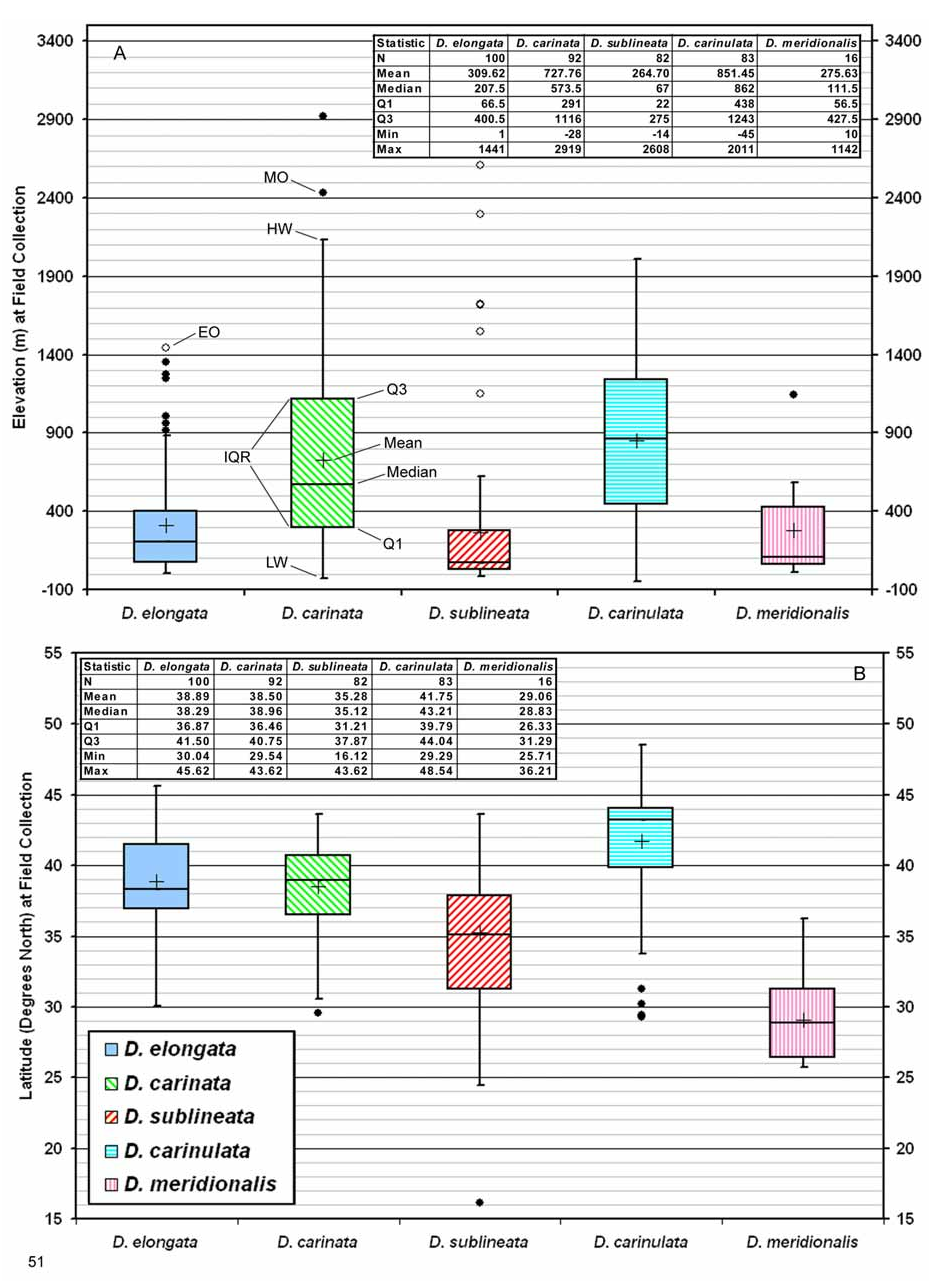
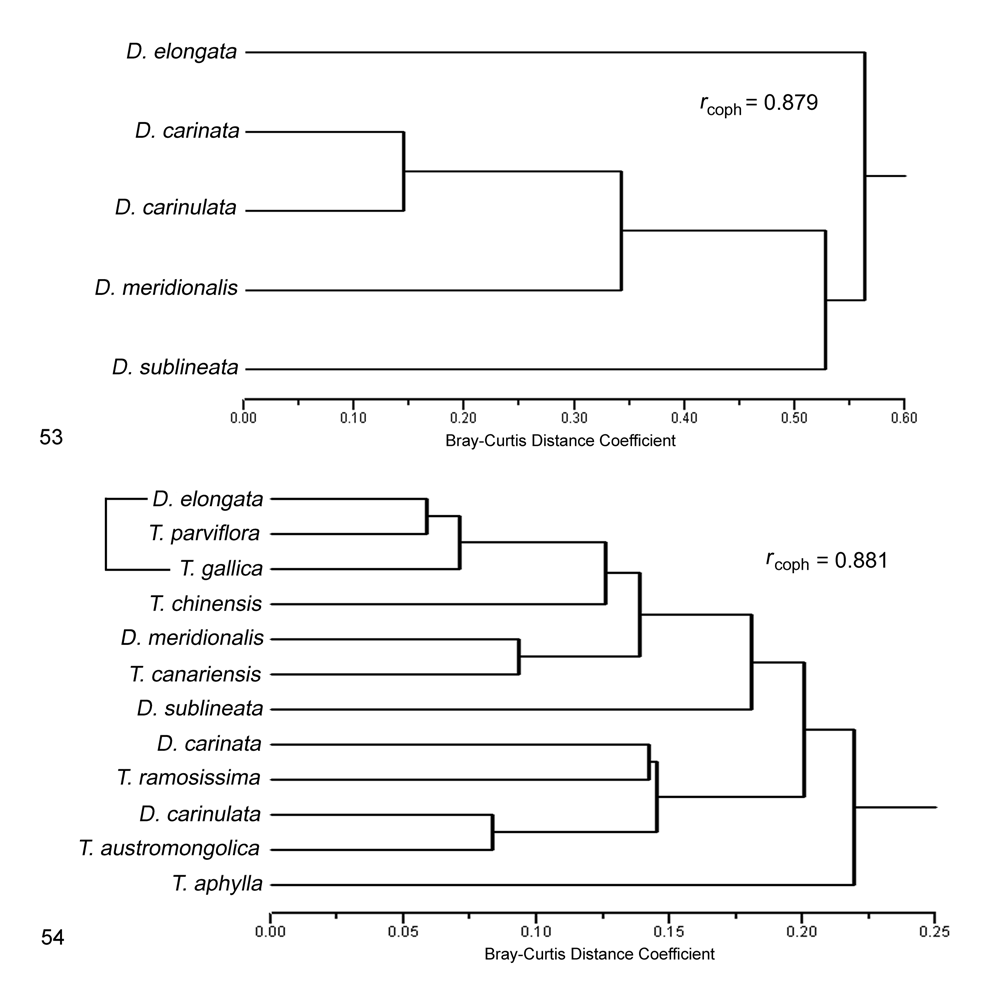
Biogeography. Comparative. Diorhabda meridionalis differs from other tamarisk beetles by the following combination of biogeographic characteristics: (1) primarily maritime and found within 300 km of the ocean at elevations usually under 600 m (ranges to 1,100 m); (2) usually found in subtropical desert and temperate broadleaf and mixed forest biomes; and (3) latitudinal range of 26– 36°N and most common at 26– 31°N. Diorhabda meridionalis is partially sympatric with D. carinata , their ranges overlapping in western Iran and Syria where they appear to be syntopic (see above discussion under D. carinata — Biogeography) (Maps 1 and 6; Table 8). Diorhabda meridionalis is parapatric with D. carinulata in southern Iran (Map 6). Diorhabda meridionalis is most similar to the species pair D. carinata / D. carinulata in terms of biomes inhabited ( Tables 9 and 10, Fig. 53 View FIGURES 53–54 ). Both D. carinata and D. carinulata differ from D. meridionalis in being primarily continental in distribution and more common north of 31°N. Diorhabda meridionalis is allopatric with D. elongata and D. sublineata (Map 1, Table 8). Diorhabda elongata and D. sublineata differ from D. meridionalis in their greater preference of the Mediterranean biome and their commonness north of 31°N. Further collections might reveal that D. meridionalis and D. sublineata are parapatric or marginally sympatric in the eastern Saudi Arabian peninsula.
Descriptive. Diorhabda meridionalis is a south central Palearctic species that may also occur at the northern tip of the Afrotropical realm in the United Arab Emirates (Map 1). It is most commonly collected from 26– 31°N in the Deserts and Xeric Shrublands biome of southern Iran in the South Iran Nubo-Sindian Desert and Semi-Desert ecoregion ( 20–390 m) (Map 6). According to the biogeographic classification of Morrone (2002), more than half of this ecoregion falls within the northwestern tip of the Oriental region (= Indo-Malayan realm of Olson and Dinerstein [2002]) which is part of the Holotropical kingdom. But we follow Olson and Dinerstein (2002), whose placement of the western border of the Indo-Malayan and Palearctic realms near the border of Pakistan and India (Map 1) is supported by borders of the corresponding MAP 1. Native distribution of the Diorhabda elongata species group. Some symbols for Diorhabda spp. overlap and some locations are approximate (see Maps 2–5).
MAP 2. Native distribution of Diorhabda elongata with occurrences of selected Tamarix spp. MAP 3. Native distribution of Diorhabda carinata with occurrences of selected Tamarix spp. MAP 4. Native distribution of Diorhabda sublineata with occurrences of selected Tamarix spp. MAP 5. Native distribution of Diorhabda carinulata and D. meridionalis with occurrences of selected Tamarix spp. MAP 6. Native distribution of the Diorhabda elongata species group in part of southwest Asia with occurrences of selected Tamarix spp. Some symbols for Diorhabda spp. overlap and some locations are approximate (see Maps 2–5).
MAP 7. Distribution of the Diorhabda elongata species group where introduced in North America with occurrences of selected Tamarix spp. Some symbols for Diorhabda and Tamarix spp. overlap.
MAP 8. Hand-fitted habitat suitability index (HSI) models for the Diorhabda elongata species group over their native distribution: A— Diorhabda elongata , B— D. carinata , C— D. sublineata .
MAP 9. Hand-fitted habitat suitability index (HSI) models for the Diorhabda elongata species group over their native distribution: A— Diorhabda carinulata , B— D. meridionalis .
faunistic kingdoms of Bobrov (1997). The distribution of D. meridionalis in the South Iran Nubo-Sindian Desert and Semi-Desert ecoregion corresponds to the western distribution of the suspected host T. dioica , and further surveys might reveal more overlap between D. meridionalis further east and north along the distribution of T. dioica in Pakistan and India (Map 5). Habitats of collection sites for D. meridionalis in southern Iran are primarily river banks, but also include saline marshes, coastal savannas, clay semi-deserts, and weedy fields ( Hoberlandt 1981, 1983). Diorhabda meridionalis is also found in the Mediterranean Forests, Woodlands and Scrub biome in Syria ( 36°N, ca. 380 m), the Temperate Broadleaf and Mixed Forests biome of western Iran ( 29°N; ca. 60–500 m) and the Montane Grasslands and Shrublands of southern Iran ( 27°N) to ca. 1,102 m elevation at Sekand ( Table 9). Most collection sites are within 100 km of the sea, the maximum distance being 309 km at Jolow Gir, Iran ( Table 7; Map 5).
MAP 10. Hand-fitted habitat suitability index (HSI) models for the Diorhabda elongata species group over western North America: A— Diorhabda elongata , B— D. carinata , C— D. sublineata (See Maps 7 or 13 for legend of symbols for sites of Diorhabda establishment, release, or planned release).
MAP 11. Hand-fitted habitat suitability index (HSI) models for the Diorhabda elongata species group over western North America: A— Diorhabda carinulata , B– D. meridionalis (See Maps 7 or 13 for legend of symbols for sites of Diorhabda establishment, release, or planned release).
MAP 12. Composite map of habitat suitability index (HSI) models scoring among the top 15% among the Diorhabda elongata species group over their native distribution. The composite map depicts the estimated most suitable Diorhabda species or group of species for a given area, not the total potential range for any given species. Some symbols for Diorhabda spp. overlap and some locations are approximate (see Maps 2–5).
MAP 13. Composite map of habitat suitability index (HSI) models scoring among the top 15% among the Diorhabda elongata species group over North America. The composite maps depict the estimated most suitable Diorhabda species or group of species for a given area, not the total potential range for any given species. Some symbols for Diorhabda spp. overlap.
Potential in Tamarisk Biological Control. Summary. The southern tamarisk beetle may potentially be the best suited Diorhabda species for biological control of tamarisk in portions of maritime subtropical deserts such as the Sonoran Desert and Tamaulipan Mezquital (Map 13). Overseas collections of D. meridionalis may be obtained from the South Iran Nubo-Sindian Desert and Semi-Desert in southern Iran and southwest Pakistan (Map 6). The preference of D. meridionalis for T. ramosissima / T. chinensis versus T. aphylla will probably be a critical issue for host range testing. In addition, the potential effects of D. meridionalis on Frankenia spp. are not known.
Discussion. We find no reports of damage to tamarisk by the southern tamarisk beetle. This species can be abundant in areas such as Rask, Iran from which we examined a large series of at least 159 beetles collected in 1973 by the Czechoslovak-Iranian entomological expedition in an area where Hoberlandt (1981) recorded the presence of T. dioica . Diorhabda meridionalis probably attacks several Tamarix spp. , as do other species in the D. elongata group, and it may also attack Tamarix introduced into North America. Host range studies are needed to verify that D. meridionalis is restricted in host range to Tamarix spp. and determine its preference for various Tamarix spp. introduced into North America. The Palearctic annual herb Frankenia pulverulenta , a close relative of Tamarix found at a collection site with D. meridionalis in Iran (see above), should be included in host range studies. Frankenia pulverulenta is naturalized in the U.S. states of Utah and Oregon ( Welsh et al. 1987, USDA Natural Resources Conservation Service 2008).
Maritime subtropical Deserts and Xeric Shrublands from 26– 31°N may be most suitable for D. meridionalis in North America (Maps 5–6). Suitable North American ecoregions include the southern Sonoran Desert , the eastern Tamaulipan Mezquital , the Baja California Desert, and the Gulf of California Xeric Scrub (Map 13).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Diorhabda meridionalis Berti & Rapilly, 1973
| Tracy, James L. & Robbins, Thomas O. 2009 |
Diorhabda carinulata meridionalis
| Warchalowski, A. 2003: 328 |
| Berti, N. & Rapilly, M. 1973: 881 |