
Diorhabda elongata ( Brullé, 1832 )
|
publication ID |
https://doi.org/10.11646/zootaxa.2101.1.1 |
|
persistent identifier |
https://treatment.plazi.org/id/038C2C5B-AC54-FF85-3B91-FB19F290D0D2 |
|
treatment provided by |
Felipe |
|
scientific name |
Diorhabda elongata ( Brullé, 1832 ) |
| status |
|
Diorhabda elongata ( Brullé, 1832)
Mediterranean tamarisk beetle
( Figs. 1, 2 View FIGURES 1–9 , 10–13 View FIGURES 10–13 , 14 View FIGURES 14–18 , 19 View FIGURES 19–23 , 24 View FIGURES 24–28 , 29 View FIGURES 29–33 , 34 View FIGURES 34–38 , 39 View FIGURES 39–43 , 44 View FIGURES 44–47 )
Galeruca elongata Brullé, 1832:271 ( Type locality: Morée [ Pelopónnisos peninsula, Greece]; Reiche and Saulcy, 1858:42 (part, France, Italy, Greece, Turkey, Syria, Lebanon [then under Syria], Egypt, Algeria, as Galleruca ); Joannis, 1866:83 (part, Greece, Cilicia [ Turkey], Lebanon [then under Syria], as Galleruca ).
Galeruca costalis Mulsant, Mulsant and Wachanru, 1852:176 ( Type locality: Cilicia [in southwest Turkey], as Galleruca ); Reiche and Saulcy, 1858:42 (established synonymy, as Galleruca ); Wilcox, 1971:63 (world catalog).
Diorhabda elongata: Weise, 1883:316 (part); 1893:635 (part; Italy, coastal env. Trieste), 1893:1132 (part, taxonomy), 1924:78 (part, world catalog, coastlands of Mediterranean Sea View in CoL ), 1925:225 (part, Egypt, area of Mediterranean Sea View in CoL ); Heyden et al., 1891:375 (part, catalog for Europe and Caucasus, southern Europe); Bedel, 1892:158 (part, France, as Dirrhabda); Winkler, 1924 –1932 (part, Palearctic catalog, Mediterranean region); Corréa de Barros, 1924:9 (part, Portugal); Peyerimhoff, 1926:359 (part, Algeria, hosts); Laboissière, 1934:53 (part, France); Porta, 1934:317 (taxonomic keys, Italy); Ogloblin, 1936:79 (part, Italy [Trieste], Greece, Turkey, Syria, Lebanon [ Palestine], S. Russia); Kerville, 1939:107 ( Turkey, env. Smyrna); Hopkins and Carruth, 1954:1129 (part, Spain, host); Pavlovskii and Shtakelberg, 1955:566 (part, Mediterranean region, southwest Russia, as Diorrhabda); Torres Sala, 1962:327 (part, Comunidad Valenciana, Spain); Lopatin, 1967:441 ( Lebanon, dunes south of Beirut); Tomov, 1969:181 ( Bulgaria), 1979:165 (biology, Bulgaria), 1984:377 (part, Turkey); Jolivet, 1967:331 (part, Mediterranean, hosts); Zocchi, 1971:86 (part, Italy); Wilcox, 1971:63 (part, world catalog); Gerling and Kugler, 1973:20 ( Israel [sic; should read Turkey]); Warchalowski, 1974:509 ( Bulgaria, as Diorrhabda), 2003:328 (part, taxonomic keys, Mediterranean region, Caucasus); Tomov and Gruev, 1975:146 ( Turkey); Georghiou, 1977:47 ( Cyprus ); Lundberg et al., 1987b:126 (Sicily); Petitpierre, 1988:93 (part, Spain); Regalin, 1997:69 (Crete); Gruev and Tomov, 1986:103 ( Bulgaria), 1998:70 (part, Bulgaria, Mediterranean); Biondi et al., 1995:12 ( Italy); Kovalev, 1995:78 (part, southwest Palaearctic); Campobasso et al., 1999:145 (part, host, Europe and Middle East); Lair and Eberts, 2001:1 (introduction plans, north Texas); Aslan et al. 2000:30 (part, Turkey); Anonymous, 2001:52 N (part, southern Europe); Chatenet, 2002:223 (part, Italy, France, Spain, Algeria); DeLoach et al., 2003a:229 (part, Greece), 2003b:126 (part, host range, ecology, Italy, Crete, Turkey), (2008, in prep.) (part, Greece); Gök and Çilbiroğlu 2003:66 ( Turkey), 2005:14 ( Turkey); Milbrath et al., 2003:225 (part, Greece); Riley et al., 2003:69,189 (part, catalog North America [introduced]); Bieṅkowski, 2004:76 (part, keys, eastern Europe [Southern Russia]); DeLoach and Carruthers, 2004a:13, 2004b:311 (part, Greece); Gök and Duran, 2004:17 (part, Turkey); Lopatin et al., 2004:127 (part, Mediterranean); Dudley, 2005a:13, 2005b:42 N (part, biological control, ex: Greece); Carruthers et al. 2006:71, 2008:262 (part, biological control, ex: Greece); Herr et al. 2006:148 (host specificity, Crete); Milbrath and DeLoach, 2006a:32, 2006b:1379 (part, host specificity, Crete); Milbrath et al., 2007 (part, host specificity, biology, Crete); Dudley et al., 2006:137 (releases in California, Crete); Everitt et al. (2007) (remote sensing, Texas, Crete); Hudgeons et al., 2007a:157 (establishment in Texas, Crete), 2007b:215 (tamarisk damage in Texas, ex: Crete); Mityaev and Jashenko, 2007:145 (biological control, ex: Greece); DeLoach (2008); Moran et al. (in press) (host range in Texas, Crete); Bean and Keller (in prep.) (diapause induction, Crete); Dalin et al. (in press) (host range; Crete); Thompson et al. (in prep.) (part, laboratory hybridization, Crete).
Diorhabda elongata ab. carinata: Porta, 1934:317 (keys, Italy).
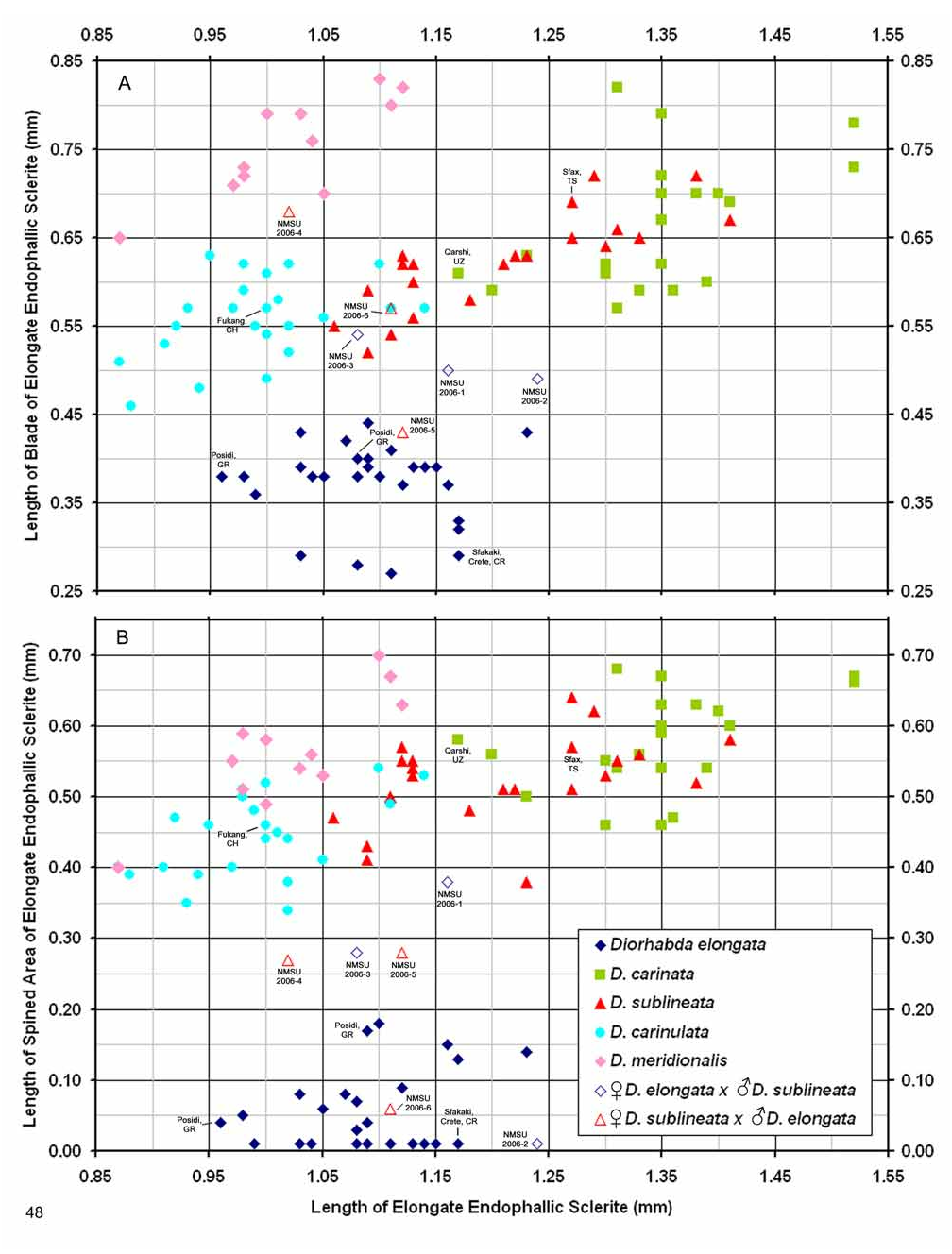
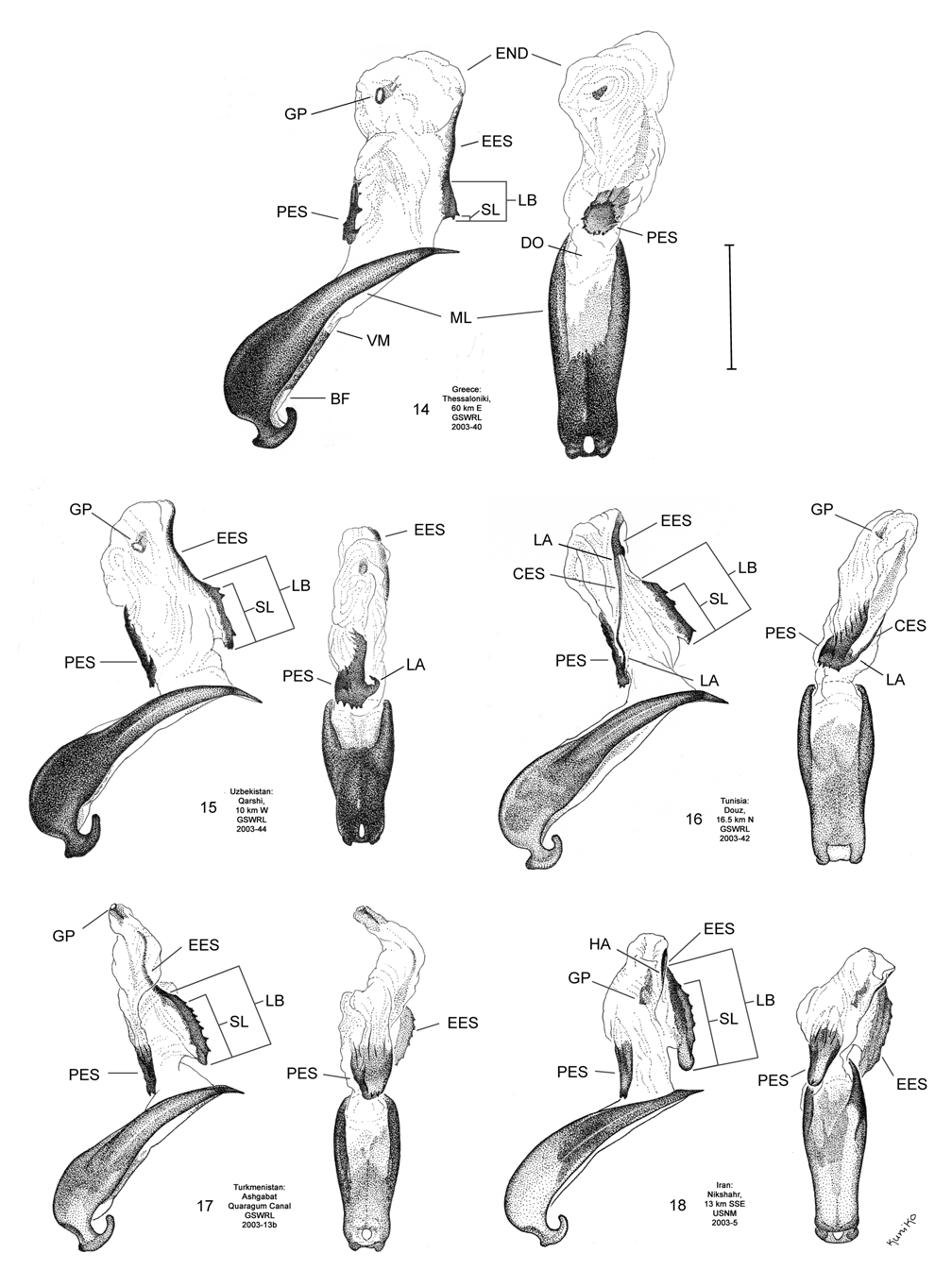
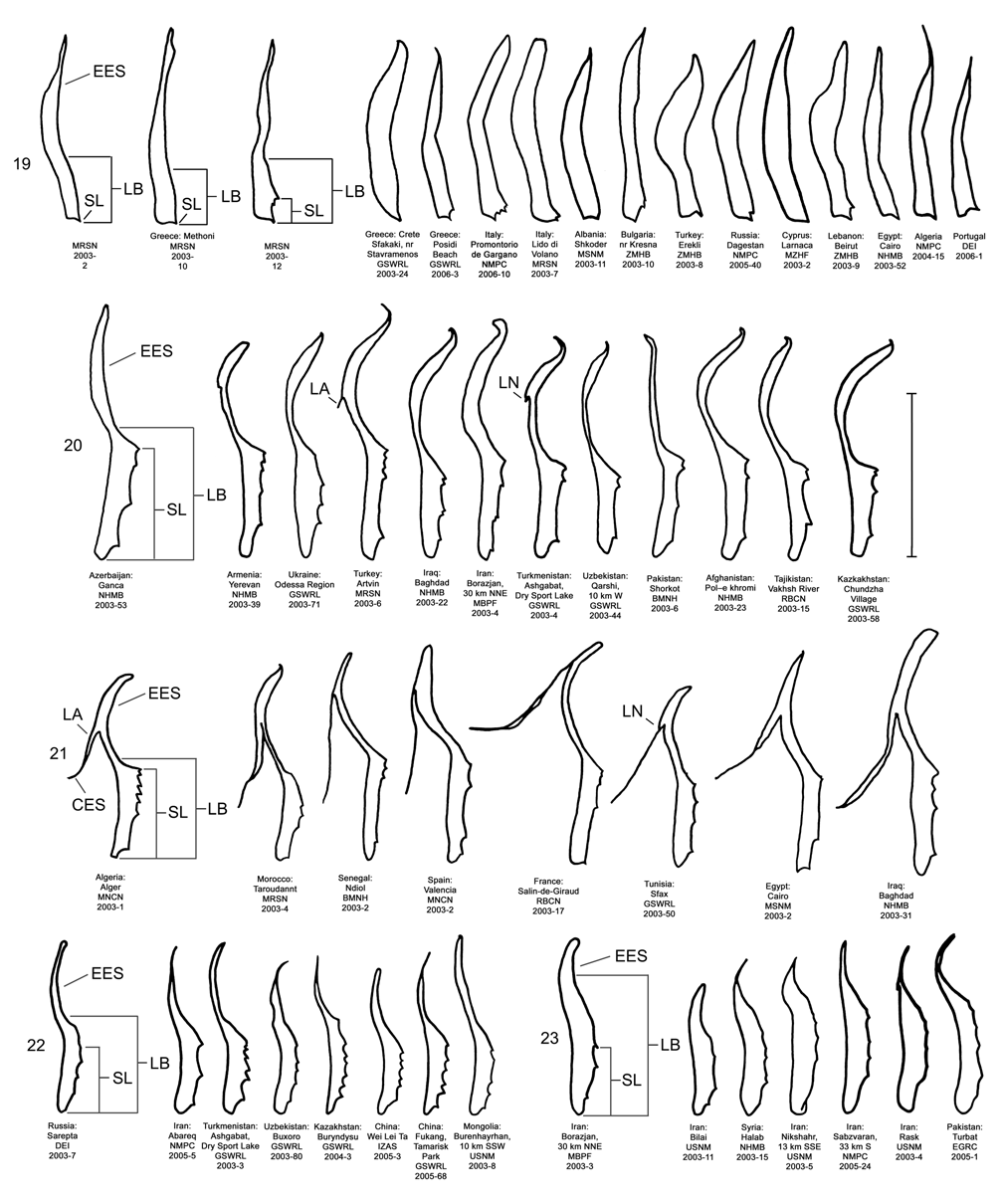
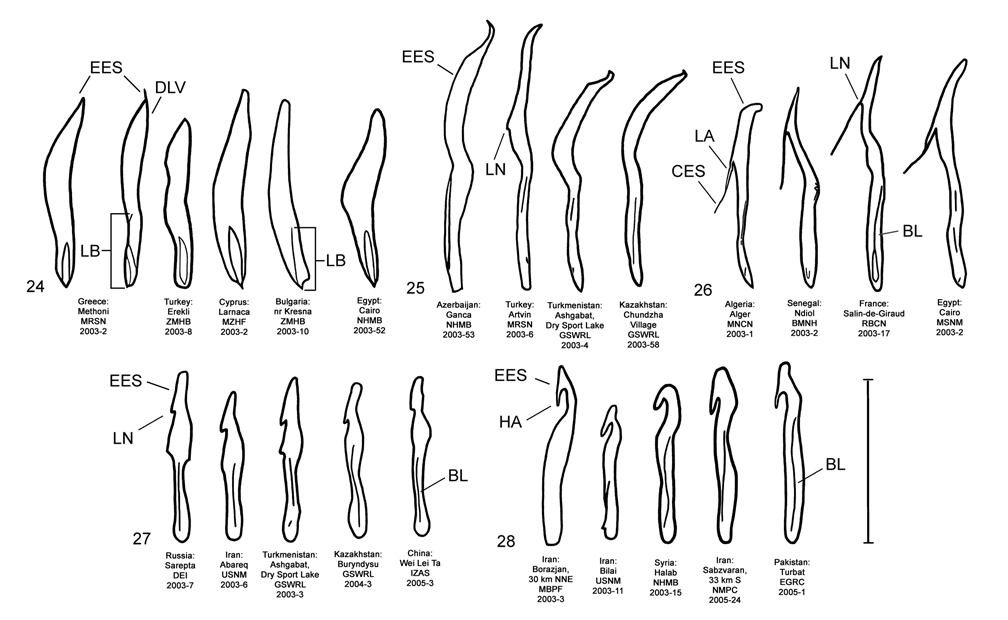
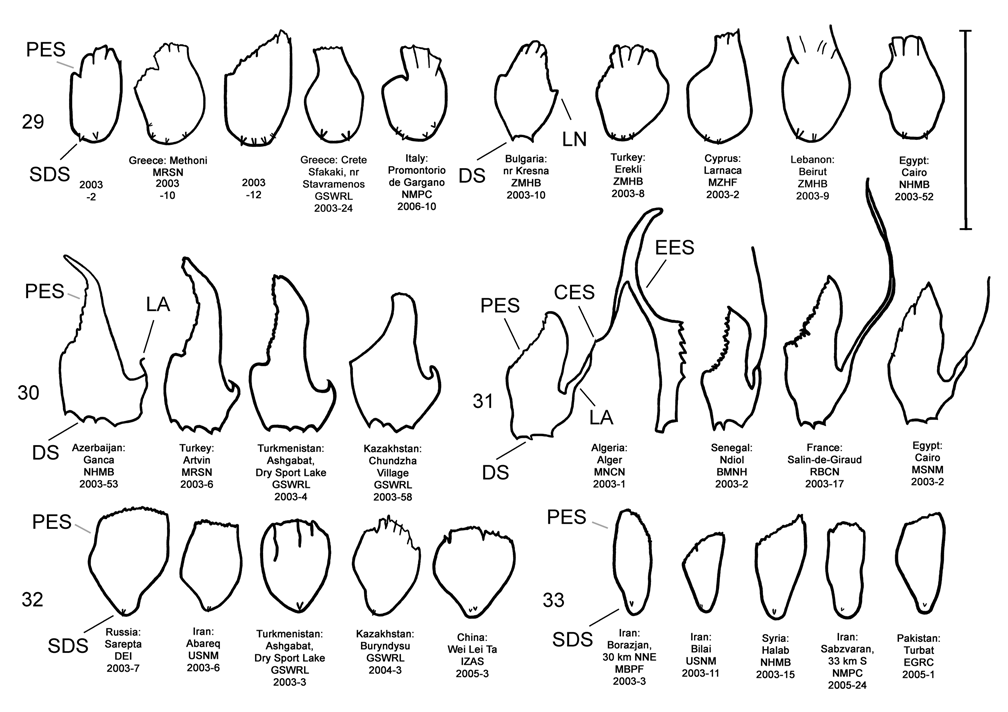
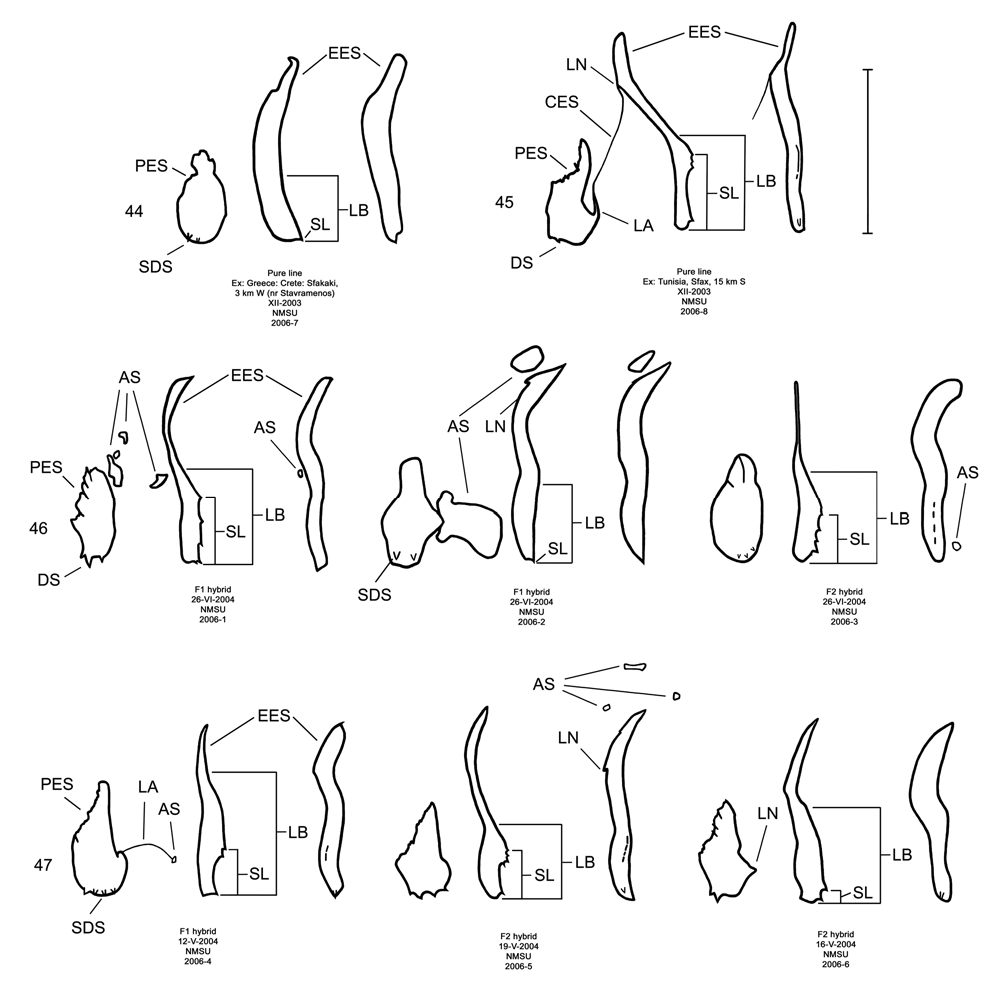
Male. Genitalia. Diorhabda elongata can be distinguished from all other members of the D. elongata group by the following combination of characters, the first two of which are unique among the tamarisk beetles: (1) the length of the spined area of the blade (SL) (armed with one to six [commonly one to three] spines) of the elongate endophallic sclerite (EES) is less than or equal to 0.16 times (or less than about one fifth) the length of the EES ( Table 3; Figs. 14 View FIGURES 14–18 , 19 View FIGURES 19–23 , 48B View FIGURE 48 ); (2) the length of the blade of the EES (LB) is less than or equal to 0.42 times the length of the EES ( Table 3; Figs. 14 View FIGURES 14–18 , 19 View FIGURES 19–23 , 24 View FIGURES 24–28 , 48A View FIGURE 48 ); (3) the EES lacks a lateral notch (pointed basally), lateral appendage or hooked apex ( Figs. 14 View FIGURES 14–18 , 19 View FIGURES 19–23 ); (4) the palmate endophallic sclerite (PES) lacks a lateral appendage, it is usually broadly rounded distally, and the one to six (commonly two to four) spines are usually subdistal with no more than two distal spines ( Figs. 14 View FIGURES 14–18 , 29 View FIGURES 29–33 ); and (5) the connecting endophallic sclerite is lacking. In the other four species of the D. elongata group, the length of the spined area of the EES (SL) (armed with three to seven spines) is greater than or equal to 0.31 times (or greater than about one third) the length of the EES ( Table 3; Figs. 20–23 View FIGURES 19–23 , 48B View FIGURE 48 ), and the length of the blade of the EES is greater than or equal to 0.43 times the length of the EES ( Table 3; Figs. 15–18 View FIGURES 14–18 , 20–23 View FIGURES 19–23 , 25–28 View FIGURES 24–28 , 48A View FIGURE 48 ). In D. sublineata , and sometimes D. carinata , the EES bears a lateral notch (pointed basally) or lateral appendage. In D. meridionalis the EES bears a hooked apex ( Figs. 25–28 View FIGURES 24–28 ). The PES always bears a lateral appendage and the distal margin is truncate serrate with usually more than two distal spines in D. carinata and D. sublineata ( Figs. 15–16 View FIGURES 14–18 , 30–31 View FIGURES 29–33 ). In D. carinulata and D. meridionalis , the distal margin of the PES differs from that of D. elongata in being narrowly or acutely rounded with one or two small subdistal spines ( Figs.17–18 View FIGURES 14–18 , 32–33 View FIGURES 29–33 ). Measurements. See Tables 2 and 3.
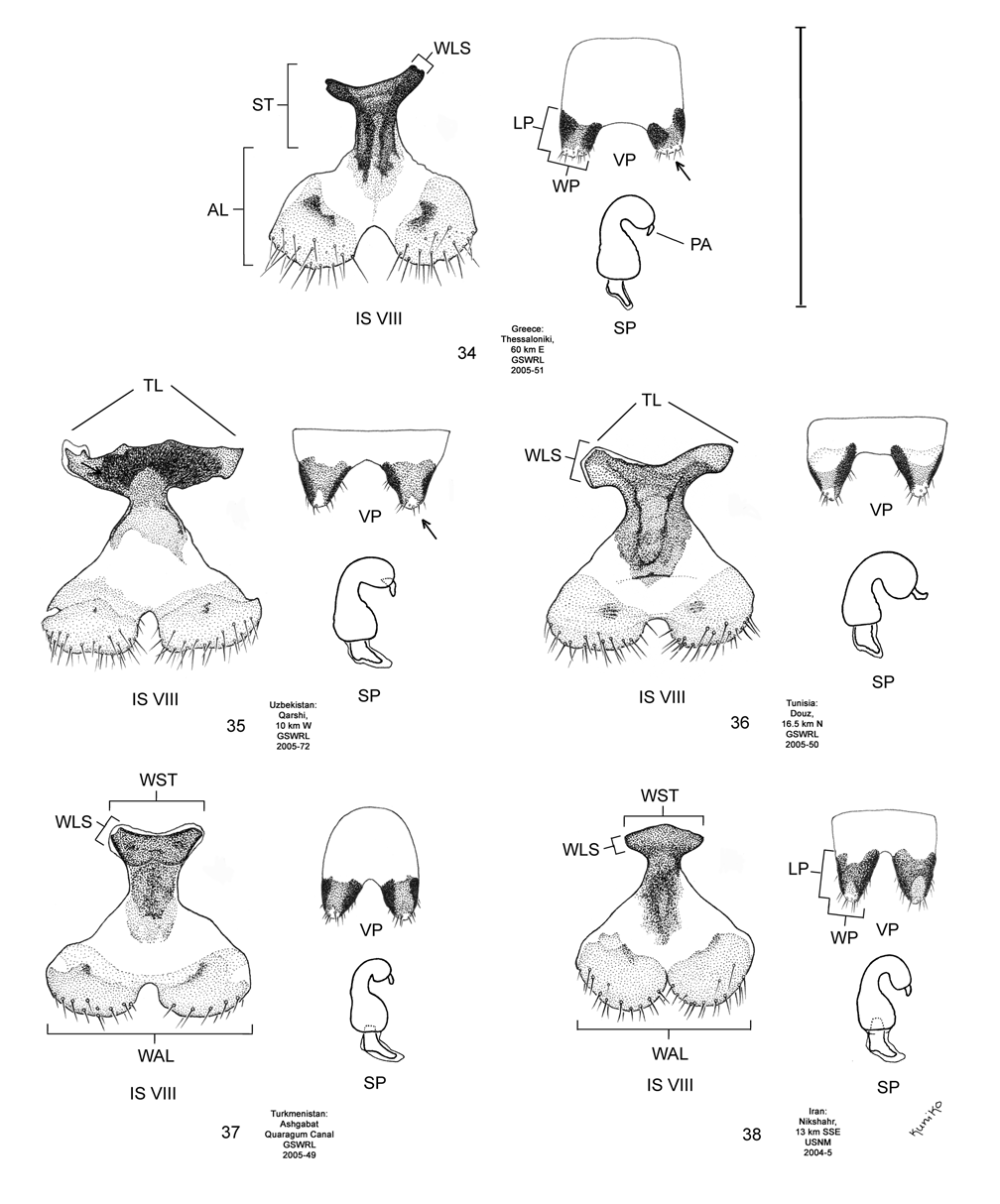
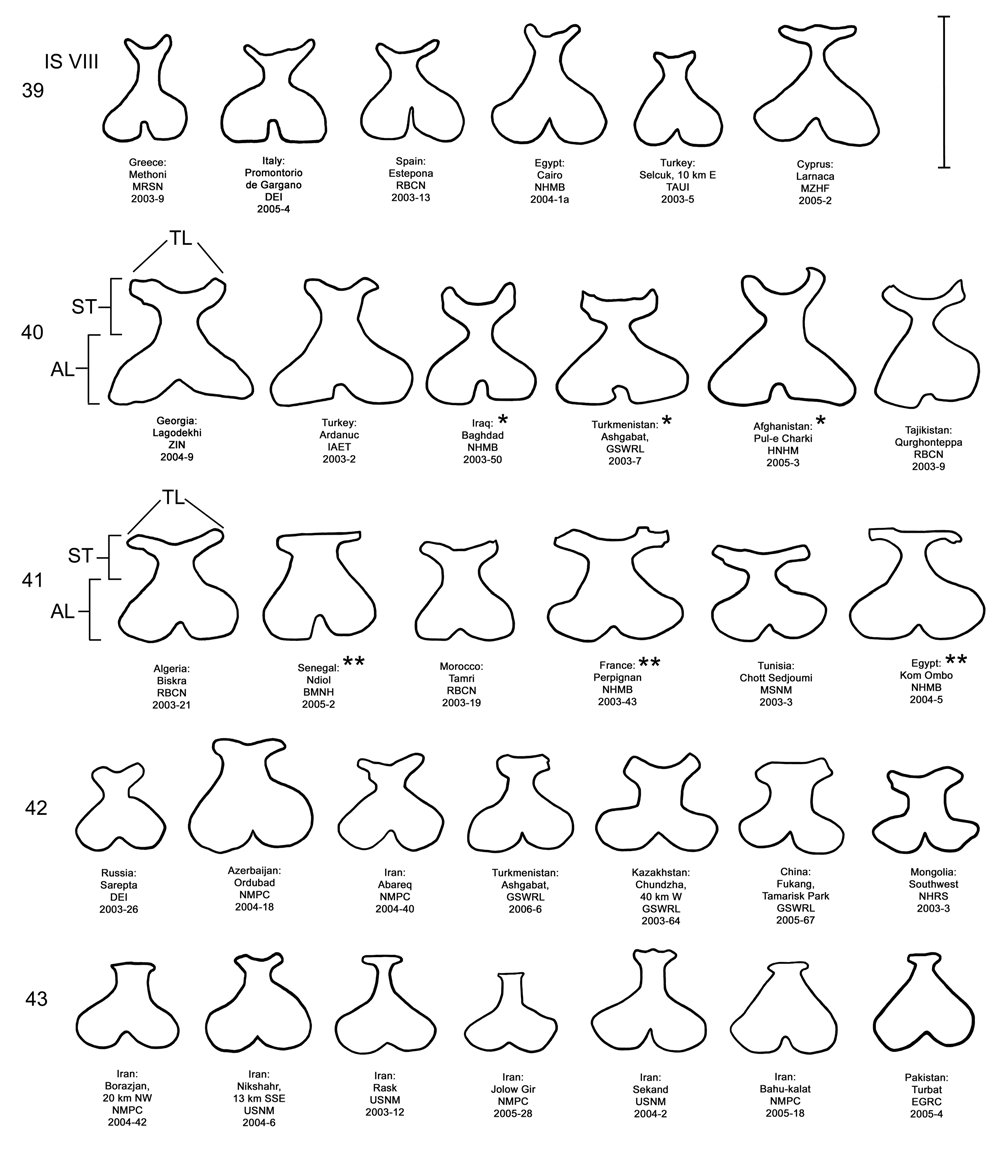
Female. Genitalia. Female D. elongata may be distinguished from all other members of the D. elongata group by the following combination of characters in the vaginal palpi (VP) and internal sternite VIII (IS VIII): (1) the vaginal palpi are both broadly rounded and wider than long with a width to length ratio (LP/WP) of 0.52–0.94 ( Fig. 34 View FIGURES 34–38 , Table 4), and (2) if the width to length ratio of the vaginal palpi is 0.94, then the width of the widest lobe of the stalk (WLS) of IS VIII is less than or equal to 0.10 mm ( Fig. 34 View FIGURES 34–38 ). In contrast to D. elongata , the vaginal palpi in D. carinulata ( Fig. 37 View FIGURES 34–38 ) and D. meridionalis ( Fig. 38 View FIGURES 34–38 ) are about as long as wide or longer with a width to length ratio of 0.94–1.36 ( Table 4). If the width to length ratio of the vaginal palpus is 0.94 in D. carinulata , then the width of the widest lobe of IS VIII is greater than or equal to 0.11 mm ( Fig. 37 View FIGURES 34–38 ). The vaginal palpi of D. carinata and D. sublineata differ in being narrowly rounded and triangulate ( Figs. 35–36 View FIGURES 34–38 ). In addition, the width of the widest lobe of the stalk of IS VIII is generally smaller in D. elongata (range 0.06–0.11 mm; Fig. 34 View FIGURES 34–38 ) compared to D. carinata (range 0.11–0.17; Fig. 35 View FIGURES 34–38 ) and D. sublineata (range 0.08–0.18 mm; Fig. 36 View FIGURES 34–38 ) ( Table 4).
Measurements. See Tables 2 and 4.
Coloration. Diorhabda elongata commonly lacks elytral vittae, but submarginal and subsutural vittae in the apical half of the elytra (evanescing toward the basal half) are not uncommon ( Fig. 1 View FIGURES 1–9 ). In all other members of the D. elongata group, elytral vittae, when present, often extend into the basal half of the elytra ( Figs. 5, 9 View FIGURES 1–9 ). Live specimens of D. elongata possess greenish-yellow tinting along veins of the elytra, probably as a result of yellow hemolymph as seen in live inflated endophalli, and this gives adults an overall olivaceous hue ( Fig. 2 View FIGURES 1–9 ). Living D. carinata have a lesser degree of greenish-yellow tinting ( Fig. 4 View FIGURES 1–9 ). Living D. sublineata ( Fig. 6 View FIGURES 1–9 ) and D. carinulata ( Fig. 8 View FIGURES 1–9 ) lack greenish-yellow tinting, probably as a result of white hemolymph seen in live inflated endophalli (also seen in D. carinata ), and they appear more tannish-yellow in hue.
Type material. Brullé’s collection from his expedition to Morée (= Pelopónnisos peninsula, Greece), which should include the type specimen(s) for D. elongata , was deposited in the Muséum National d'Histoire Naturelle in Paris, France ( MNHN) ( Groll 2006) . A curator at MNHN communicated intent to inform us of the status of the type material and perhaps lend it for examination, but after four years, we have not heard of the status of the Brullé type material. Once it can be ascertained that the type material is lost, a neotype should be designated using a dissected male specimen from the type locality of Pelopónnisos, Greece . We studied the original description by Brullé (1832), based on an unspecified number of specimens, with the included color habitus illustration, and topotypes from the Pelopónnisos peninsula, Greece .
The location of type specimens of Galeruca costalis Mulsant (Mulsant and Wachanru 1852) is uncertain. Remaining portions of E. Mulsant’s collection “Natural History of the Coleoptera of France ” should be located at the l’Institution Sainte– Marie de Saint –Chamond ( Loire), France ( Groll 2006) . We studied the original descriptions of G. costalis by Mulsant (Mulsant and Wachanru 1852) and topotypes from the Cilicia region of Turkey .
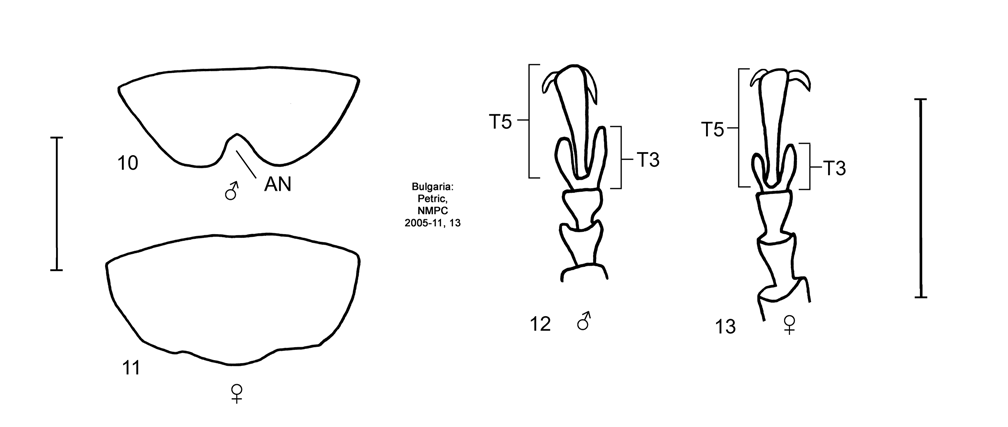
Material examined. 157♂♂ dissected (diss.), 85♀♀ diss., 127♂♂, 164♀♀. ALBANIA: 1♂ diss., Durazzo [ 41.32305°N, 19.44138°E], Lona-Rav., Diorhabda elongata det. Burlini 1967, MSNM [2003-08]; 1♂ diss., 1♂, 1♀, Elbasan [ 41.11250°N, 20.08222°E], D. elongata, USNM [2003-10]; 1♂ diss., Medua [Shengjin, San Giovanni di Medva; 41.81366°N, 19.5939°E], Matzenaver, D. elongata det. V. Zonfal, NMPC [2004-51]; 1♂ diss., Srutari [ Shkoder; 42.0675°N, 19.5131°E], D. elongata det. Burlini 1967, MSNM [2003- 11]; 1♀ diss., Scutari [ Shkoder], Heyrevsky, NMPC [2006-06]; 1♂ diss., 1♀, Valona [ Vlore; 40.46666°N, 19.48972°E], [19]08, H. Hopp, ZHMB [2003-04]; ALGERIA: 1♂ diss., [specific locality not given], Reitter, NMPC [2004-15]; BOSNIA & HERZEGOVINA: 1♂ diss., Veleś Planina [Velez Planina; 43.33°N, 18.0044°E], Herzegowina, 1879, Reitter, D. elongata, HNHM [2003-05]; BULGARIA: 1♂ diss., 1♂, 1♀, Burgas [ 42.5°N, 27.4667°E], 16-VII-1975, Vaśárhelyi, D. elongata det. V. Tomov 1983, HNHM [2003-06]; 1♀ diss., Gara Pirin [Kresna; 41.7333°N, 23.1500°E], Kresn. defile [Kresnenska Klisura or Kresna defile (gorge)], B. Kouřil, Diorhabda det. B. Kouřil, P9/46/62, NMPC [2005-25]; 1♂ diss., 1♀, Gara Sandanski [Sandanski; 41.5667°N, 23.2833°E], Struma [river], 20-VII-1956, L. Hoberlandt, NMPC [2006-04]; 1♂ diss., Kresna [ 41.7333°N, 23.1500°E], environs, Struma [river], 29-V-1984, U. Göllner, D. elongata det. V. Tomov, ZMHB [2003-10]; 1♂ diss., 1♂, 1♀, Kresnicko Def. [Kresnenska Klisura or Kresna defile (gorge); 41.8°N, 23.16667°E], VII-1932, Mac, Mař et Táb, NMPC [2006-05]; 1♀ diss., Kritschin [Krichim; 42.05°N, 24.4667°E], NHMB [2003-41]; 1♂ diss., Melnik [ 41.5167°E, 23.4°E], 23-V-1973, St. Gruszka & A. Warchalowski, GSWRL [2003-41]; 2♂♂ diss., 1♀ diss., Petric [ 42.6°N, 24.01667°E], 1932, D. Putkynĕ, D. elongata det. Sterba, NMPC [2004-24, 2005-11, 13]; 1♀ diss., Pomorie [ 42.55°N, 27.65°E], 9-V-1970, B. Gruev, D. elongata det. V. Tomov 1997, GSWRL [2003-46]; 1♂ diss., 3♂♂, 2♀♀, Sandanski [ 41.5667°N, 23.2833°E], Meridion., 10-VIII-1974, A. Warchalowski, GSWRL [2003-34, 1♂ diss.], DEI; 1♂ diss., 4♂♂, 4♀♀, Keretschkoi [Kirechdzhi Khask'oy is Sladun; 41.85°N, 26.4667°E], Macedonia, A. Schatzmayer, D. elongata det. Fleischer, DEI [2003-18]; CROATIA: 1♂ diss., Metkovic [Metkovi; 43.054167°N, 17.648333°E], Dalmatien, E.A. Bottcher (Berlin), 168, MZHF [2005-03]; 1♂ diss., Metkovich [Metkovi], Dalmatia, NHMB [2003-25]; 1♀ diss., 1♀, Opuzen [ 43.00972°N, 17.56555°E], 26-V-1974, G.J. Slob, RBCN [2003-07], ZMAN; 1♂ diss., 5♂♂, 10♀♀, Ušce Neretve D. [ 43.01917°N, 17.44389°E; Neretva river mouth], 8-IX-1948, Novak [ ZMHB], D. elongata det. Novak, HNHM [2003-04], ZMHB [ 2♂♂, 4♀♀]; CYPRUS : 1♂ diss., [specific locality not given], 1876, L. Shrader, Coll. Kunnemann, DEI [2003-13]; 2♂♂ diss., 1♀ diss., 6♂♂, 5♀♀, Larnaka [ Larnaca; 34.916667°N, 33.6333°E], 25-VI to 1-VII-1939, Håkan Lindb., D. elongata det. Harald Lindb. [ 3♂♂, NMPC; 1♂, 1♀, NHRS], MZHF [2003-02, 2005-01, 02], NMPC, NHRS; 1♂, 1♀, Larnaca, Kamares Aqueduct, 19-X-1932, A. Ball, D. elongata det. V. Laboissière 1939, IRSNB; 2♂♂ diss., 1♀ diss., 3♀♀, Mt. Arménien [ 35.2875°N, 33.525°E], NMPC [2004-02, 2005-01, 02]; EGYPT: 3♂♂ diss., 2♀♀ diss., 1♀, Cairo [ Al Qahirah; 30.05°N, 31.25°E], NHMB [2003-16, 52, 2004-01a, 2005-02, 03]; GREECE: 1♂ diss., Agia Marina, Star Beach [ 35.5193°N, 23.9275°E], Crete, 10-IX-2001, R. & N. Carruthers, on Tamarix sp. foliage, EIWRU-2001-1016, USNM [2003-18]; 1♀ diss., 3♀♀, Ammos [ 36.91270°N, 27.28170°E], Kos [Island], 27-VIII to 1-IX 1981, T. Palm, MZLU [2005-08]; 6♂♂ diss., 2♀♀ diss., Astro Beach [ 37.4433°N, 22.74861°E], north of, Pelopónnisos, 25-V-2003, J. Kashefi, on T. hampeana, GSWRL [2004-10, 2005-38, 39, 41–45]; 1♀ diss., Athens environs [ 37.96138°N, 23.63888°E], Dr. Krüper, NHMB [2004-03]; 1♂, 2♀♀, b. Athens, v. O., Collectie C. et O. Vogt Acq. 1960, ZMAN; 1♂ diss., 1♂, 2♀♀, Athos [ 40.166667°N, 24.333333°E], Macedonia, Schatzmayer, Coll. V. Heyden, D. elongata det. Fleischer, DEI [2003-19]; 2♂♂ diss., 3♂♂, Athos, Macedonien, 1908, Schatzm., NHMB [2003-26, 2006-02]; 1♀ diss., 1♂, 4♀♀, Attica [ Attiki; 38.083333°N, 23.5°E], Stussiner, O. Leonard, DEI [2003-15]; 1♂, 3♀♀, Attica, Reitter, Coll[ection] Kambersky, D. elongata, NMPC ; 2♂♂ diss., 1♂, 2♀♀, Attica, Reitter, 269 [ MZLU ♀] D. elongata Brulle Emm. Reitter [ ZMAN; 1♂, 1♀], Collectie P.v.d. Wiel Acq. 1962 [ ZMAN], MZLU, ZMAN [2008-05, 17], ZMHB [ 1♂]; 1♂, Attica, Witte, coll[ection] V. Heyden, DEI; 1♀, Attica, Hymettos [Mts.], Emge., ZMAN; 1♀, Attica, v. Oertzen, ZMAN; 1♂, Attika, ZMHB; 1♀ diss., 1♂, 1♀, Crete [specific locality not given], D. elongata det. K. Lopatin, HNHM [2005-01]; 1♀ diss., 1♀, Diakophto [Diakofto; 38.1795°N, 22.2001°E], Pelop., IV-1936, Mař et Táb., Coll[ection] Bartoň, NMPC [2004-34]; 1♀ diss., Dimilia [ 36.2834°N, 28.0085°E], Rhodes [island], 16-VII-1933, A. Mochi, D. elongata det. Burlini 1967, MSNM [2003-14]; 1♂ diss., Euboea [Evvia Island; specific locality not given; approx. loc.: 38.5200°N, 23.72000°E], Coll. Kraatz, DEI [2003-14]; 1♂ diss., 1♀, Fileremo [Filerimos Hill; 36.3985°N, 28.1382°E], Rodi [Rhodes], 22-IV-1932, A. Schatzmayer, D. elongata det. Burlini 1967, MSNM [2003-12]; 1♀ diss., Fileremo [Filerimos Hill], Rodi [Rhodes], 10-VII-1933, A. Mochi, D. elongata det. Burlini 1967, MSNM [2008-02]; 1♂ diss., Heraklion [Irakleion; 35.325°N, 25.1306°E], Crete, 13-IX-2001, R. & N. Carruthers, lab reared from eggs collected from Tamarix sp. foliage, EIWRU-2001-1016, USNM [2003-09]; 1♀ diss., 1♀, Heraklion [Irakleion], 18-X-2001, R & N. Carruthers, lab reared from eggs on Tamarix spp. [at Albany, CA], USNM [2005-01]; 1♀ diss., 1♂, 1♀, Heraklyon [Irakleion], Kreta, 22-V-1995, J. Blümel, D. elongata det. Erber 2001, ZMHB [2008-04]; 1♀ diss., 2♂♂, 1♀, Hypati [Ypati; 38.88590°N, 22.23350°E], Pfeffer, D. elongata det. V. Tomov 1984 [ 1♂ diss.], NMPC [2004-46]; 1♂ diss., 1♂, 3♀♀, Hypati [Ypati], IV-1936, Mař et Táb., Coll[ection] Bartoň, NMPC [2006-07]; 1♂ diss., 6♂♂, 3♀♀, Ilis [ 37.8833°N, 21.3833°E], Olympia, 2–3-X- 1962, Ent. Exc.Zoöl Mus., ZMAN [2008-14]; 1♂ diss., 2♂♂, 3♀♀, Ixia [Ixos; 36.4167°N, 28.18330°E], Rhodes, 19-IV to 1-V-1973, A. Teunissen, RBCN [2003-04], ZMAN [ 1♀]; 1♂ diss., Kalamata [ 37.0389°N, 22.1142°E], 22-IX-1982, Roland Bö, 579, D. elongata det. Eber 1989, ZMHB [2008-01]; 1♂ diss., Karia environs [ 39.98330°N, 22.4000°E], Olymp[us Mt.], 1000 m [elev.], 30-VI-1980, Probdt, D. elongata det. V. Tomov 1983, NMPC [2005-26]; 1♂ diss., 4♂♂ Kardamena [Kardamaina; 36.7814°N, 27.1425°E], Kos [Island], 2–6-IX-1981, T. Palm, MZLU [2005-01]; 2♀♀ diss., 1♂, 5♀♀, Karpathos Town [ 35.5°N, 27.23333°E], Ormos Pigadia env., 0–150 m, Karpathos Island, 28-VI to 3-VII-1996, W.T. Edzee, D. elongata det. A. Teunissen 1998 [ ZMAN ♀], RBCN [2003-12, 2005-01; 2♀♀], ZMAN; 1♀ diss., Katakolo [Katakolon; 37.6500°N, 21.3167°E], Morea v. O., Collectie C. et O. Vogt Acq. 1960, ZMAN [2008-15]; 1♂ diss., 2♂♂, Kerkyra [ 39.6200°N, 19.91970°E], Korfu [Kérkyra Island], V-1964, Palm, MZLU [2005-07]; 1♂ diss., 1♀, Katafourko [Katafourkon; 38.9981°N, 21.1511°E], (Etol), 27–30-V-1998, P. Foot, ZMAN [2008-04]; 1♀ diss., 1♂, 1♀, Korfu, Insel [Kérkyra Island; specific locality not given], NHMB [2003-28] [ 1♂ resembles aberration bipustulata]; 1♂ diss., 6♀♀, Korission [Korissíon, Limni (sea); 39.44611°N, 19.90694°E], Korfu [Kérkyra Island], V-1964, Palm, MZLU [2005-06]; 1♂ diss., 1♀, Kos [Island, specific locality not given], Südl. Sporaden, Oertzen, D. elongata , 86202, ZMHB [2003-11]; 2♂♂ diss., 1♀ diss., Kyparrisia [ 37.2577°N, 22.6533°E], Pelopónnisos, 26-V-2003, J. Kashefi, on T. parviflora, GSWRL [2004-11, 23, 24]; 1♂ diss., Lixourion [ 38.201944°N, 20.431389°E], Cephalonia, 26-VIII-1899, A. Porta, D. elongata det. A. Porta, MSNM [2003-15]; 1♀ diss., Mesolongion [ 38.369167°N, 21.429167°E], NHMB [2003-40]; 1♂ diss., Malia Beach [Mália; 35.2883°N, 25.4667°E], meadow west of hotel, Kreta [ Crete island], 5–12-V-1979, R. Danielsson, D. elongata det. Mohr 1980, MZLU [2005-12]; 4♂♂ diss., 2♀♀ diss., 3♂♂, 4♀♀, Methoni [ 36.81666°N, 21.7°E], Peloponneso [ Pelopónnisos Penninsula], 23-IX-1997, MRSN [2003-02, 09, 10, 11, 12, 13]; 3♂♂, 1♀, Morea [ Pelopónnisos Penninsula], v. Oertzen ( 2♂♂, 1♀), Collectie C. et O. Vogt Acq. 1960, ZMAN; 1♂ diss., 1♂, Oeta [Mt.; specific locality not given], IV-1936, Mař et Táb., Coll[ection] Bartoň, NMPC [2004-36]; 1♂ diss., 1♂, 1♀, Molyvos [Mithymna; 39.3667°N, 26.1667°E], Lesbos [Lesvos Island], 4–11-V-1992, J.H. Woudstra, ZMAN [2008-06]; 1♂ diss., 5♂♂, 6♀♀, Olympia [Archaía Olympia; 37.6500°N, 21.6333°E], Alpheios [Alfeiós] Valley, Morea [ Pelopónnisos Penninsula], 6-V-1930, A. d'Orchymont, D. elongata det V. Laboissière 1939, IRSNB [2006-04]; 1♂ diss., Olympia [Archaía Olympia], Morea, v. Oertzen, ZMAN [2008-19]; 1♀ diss., 1♀, Pahia Ammos [Pakhia Ammos; 35.11670°N, 25.8000°E], Kreta [ Crete Island], 16-V-1975, MZLU [2005-11]; 1♀ diss., 1♂, Pantokrator [Pantokrátor Mt.; 39.75396°N, 19.86277°E], Korfu [Kérkyra Island], V-1964, Palm, MZLU [2005-09]; 1♂ diss., 2♀♀, Paradisi [ 36.4006°N, 28.0796°E], Rhodes, 21-IV-1996, A. Teunissen, RBCN [2003-18], ZMAN [ 1♀]; 1♀ diss., Peloponn. [ Pelopónnisos Penninsula], D. e. var. sublineata det. K. Lopatin 1958, HNHM [2004-06]; 1♂ diss., 1♀, Piraus [Peiraiefs] env., Athens, 2-VIII-1982, Steinhausen, D. elongata det. Steinhausen, RBCN [2003-14]; 1♀ diss., Pirgos [ 37.6833°N, 21.45°E], Peloponnese [ Pelopónnisos Penninsula], 17-VI-1986, H. Hebauer, D. elongata det. Doberl 1989, MBPF [2003-05]; 1♀ diss., 1♂, 1♀, Plimmiri [ 35.929°N, 27.8584°E], Rhodes, 20-IV-1996, A. Teunissen, RBCN [2003-20], ZMAN [ 1♂]; 1♂ diss., 1♀ diss., Possidi [Posidi Beach; 39.96467°N, 23.36483°E], Kassandra, Halkidiki, 3-VI-1989, R. Sobhian, on Tamarix sp. , “RS,89,I09”, USNM [2005-03, 04]; 3♂♂ diss., 2♀♀ diss., 27♂♂, 14♀♀, Possidi [Posidi Beach], Kassandra, 39° 57.88' N, 23° 21.89' E ( GPS), 28-IV-1999, J. Kashefi & R. Sobhian, on Tamarix spp. , 9063, 9091, 9092, 9094, 9095, 9097, 9098–9118, 9120–9124, D. elongata det. I.K. Lopatin 1999 [ 2♂♂ diss., 2♀♀ diss., 23♂♂, 8♀♀], GSWRL [2003-13, 2005-27, 28, 29, 2006-03]; 6♂♂ diss., 4♀♀ diss., Posidi Beach [not Thessaloniki, as in some shipping records; Halkidiki region], 3-X-2002, J. Kashefi, EIWRU-2002-1009, USDA/ARS lab colony at Temple, Texas, voucher [J.L. Tracy, 2002–2003], USDA/ARS lab colony at Albany, California, voucher, GSWRL [2003-17, 19, 26, 27, 30, 2005-33, 34, 35, 64, 65] [introduced in New Mexico and Texas in 2004] [used in biological studies Herr et al. (in prep.), and Bean and Keller (in prep.)]; 1♂ diss., Rhodos umg. [env.] [Rodos; 36.4408°N, 28.2225°E], 1-VIII-1982, Steinh., D. elongata R. Beenen det. 1996, ZMHB [2008-02]; 1♀, Rhodes [island, specific locality not given], Reitter, NMPC; 1♂ diss., 1♀, Rhodos [Rhodes island,], 15- VIII-1970, Rich Dahl, D. elongata det. L. Borowiec, MZLU [2005-05]; 1♀ diss., 1♀, Rhodos [Rhodes island], 24-VI to 4-VII-1958, Palm, MZLU [2005-10]; 1♀, Rhodes, Reitter, NMPC; 1♀, Salonich [Thessaloniki; 40.64027°N, 22.94388°E], Schatzmayer, coll[ection] V. Heyden, DEI; 1♂ diss., 1♀, Saloniki [Thessaloniki], Vardarebene [Vadar River], ZMHB [2003-02]; 1♂ diss., Servia, 5 km NW [ 40.2204°N, 21.9571°E], Makedonia, 19-VIII-1965, E. A. Blommers, ZMAN [2008-09]; 1♂ diss., 1♂, Sigri [Sigrion; 39.2167°N, 25.8500°E], Lesbos [Lesvos Island], 8-V-1992, J.H. Woudstra, ZMAN [2008-10]; 7♂♂ diss., 6♀♀ diss., 2♂♂, 3♀♀, Sfakaki, 3 km west [ 35.3833°N, 24.6°E; near Stavramenos, Crete], road from Irakleion to Rethymnon, 5-IV-2002, R. Carruthers & J. Kashefi, shipment EIWRU-2002-1002, USDA lab colony at Temple, Texas, voucher ( 28-VI-2002, 6-IX-2002, J.L. Tracy; 1♂ diss., 28-VIII-2002, 1♂, 2♀♀, 26- VIII-2002, T.O. Robbins; 1♂ diss., 1♀ diss, 30-VI-2003, 28-VII-2003, L. Milbrath [nos. 1278, 1274]), GSWRL [2003-08, 10, 20, 24, 25, 31, 32, 40, 46, 47, 79, 82, 83] [introduced in California, New Mexico and Texas in 2003] [used in biological studies of Milbrath and DeLoach (2006a, 2006b); Milbrath et al. (2007); Moran et al. (in press); Herr et al. (2006, in prep.); Bean and Keller (in prep.); Hudgeons et al. (2007a, 2007b); and Thompson et al. (in prep.)]; 1♀ diss., 3♀♀, Sparta [Sparti; 37.0733°N, 22.4297°E], Pelopones, 1935, Mařan et Stĕp., Coll[ection] Bartoň, NMPC [2004-32]; 1♂ diss., Taygetos [Mt.; specific locality not given], Pelopon. [ Pelopónnisos Penninsula], 1935, Dr. Purkyne, NMPC [2004-37]; 1♀ diss., Sparti, 5 km SW [ 37.0386°N, 22.3985°E], Lakonia [Laconia Prefecture], 27-IX-1962, Ent. Exe. Zoö. Mus., ZMAN [2008-08]; 1♀ diss., Thérisos [Therissos; 35.3333°N, 25.1167°E], 3 km W Iráklion [Irakleion], Kriti [ Crete], 14-X-1972, A.C. & W.N. Ellis, ZMAN [2008-07]; 4♂♂ diss., 2♀♀ diss., 1♂, 9♀♀, Thessaloniki, 60 km E [ 40.64°N, 23.48°E], 117 m [elev.], 30-IV-1999, Kashefi & Sobhian, on Tamarix spp. , 9126–9129, 9131–9136, D. elongata det I. Lopatin 1999 [ 5♂♂, 5♀♀], GSWRL [2003-40, 2005-26, 30, 51, 2006-04, 05]; 1♀ diss., Trianta [ 36.4°N, 28.16666°E], Rhodes, 29-IV-1932, A. Schatzmayer, D. elongata det. Burlini 1967, MSNM [2003-13]; 1♀ diss., Xylokastron [ 38.08333°N, 22.63333°E], 29-V-1964, E. Junger, ZMHB [2003-03]; ITALY: 1♂ diss., 1♀ diss., 2♂♂, Alcantara River banks by sea [ 37.8067°N, 15.2594°E], Sicilia [ Sicily], V- 1926, NHMB [2003-49, 2005-01]; 1♂ diss., 1♂, Ancona [ 43.6333°N, 13.5000°E], Zeller, 611, 172, DEI [2008-01]; 1♂ diss., 2♂♂, 3♀♀, Cotrone [Crotone, resembles Botrone; 39.0833°N, 17.1333°E], Calabria, 26- VIII-[18]88, A. Fiori, ZMHB [2008-06]; 1♂ diss., Fiumi di Pollina [ 38.01667°N, 14.16667°E], Sicilien [ Sicily], 1982, T. Palm, D. elongata det. Thure Palm, coll[ection] T. Palm, MZLU [2005-02]; 1♀, Lido [ 45.6783°N, 13.4025°E], O. Funk, ex. coll. J. Weise, ZMHB; 1♂ diss., Lido di Volana [Lido di Volano; 44.79638°N, 12.27083°E], Littorale Ferrarese, 24-V-1959, A. Giordani Soika, Tamarix dunes, MRSN [2003- 07]; 1♂ diss., 1♂, Messina [ 38.1833°N, 15.5667°E], Sicilia [ Sicily], Reitter, D. elongata [ NMPC], Coll. Dr. J. Erdo’s [ HNHM], HNHM [2004-07], NMPC; 1♂ diss., Messina, Fobria [?], 21-IX-1952, A. Porta, Galerucella grisescens det. Antonio Porto, MSNM [2008-01]; 4♂♂ diss., 5♀♀ diss., 2♂♂, 4♀♀, Monte Gargano [Promontorio de Gargano; 41.8333°N, 16.0°E], 30-IV to 13-V-1907, M. Hilf, Coll. O. Leonard, D. elongata [ 1♂], DEI [2003-01, 02; 2005-02, 03, 04], NMPC [2006- 10 ♂; 2 ♀♀], ZMHB [ 2♀♀]; 1♂ diss., Pescara [ 42.4667°N, 14.2167°E], Abruzzo, 12-IV-1906, A. Fiori, ZMHB [2003-05]; 1♂ diss., 1♀, Piano Torre [resort near Altavilla Milicia, Sicily; 38.0333°N, 13.5333°E], X-1908, [ ZIN 2004-12]; 1♀ diss., Pineta [ 42.45°N, 14.2333°E], Lido Bruno, Puglia Prov., 7-III-1981, F. Montemurro, seaside, D. elongata det. D. Sassi 1995, GSWRL [2003-60]; 1♂ diss., 1♀, 1♂, Pioppi ([nr.] Salerno) [ 40.1833°N, 15.0833°E], V– VI- 1965, H.K. Mohr, DEI [2003-09]; 1♀ diss., Pioppi ([nr] Salerno), 23-X-1964, W. Liebmann, DEI [2003-24]; 1♂ diss., 1♂, Rendina Val[ley] [Rendina River; approximate location: 41.0166°N, 15.7382°E], Leone [?], Basilicata, 1901, A. Fiori, ZMHB [2008-07]; 1♂ diss., Rodia [Villagio Rodia, suburb of Messina], X-1932, F. Vitale, Le Moult vend. via Reinbek Eing. 1-1957, 78, D. e. ab. carinata det. V. Laboissière, ZMUH [2006-01]; 1♂ diss., Rosalina Mare [45.1192°, 12.3133°E], 4-VII-1962, A.G. Soika, beach dune with Ammophiletum, MRSN [2003-05]; 1♀ diss., Sinnaro [Contrada de Sinnaro, quarter of Messina], 16-VI-1929, F. Vitale, Le Moult vend. via Reinbek Eing. 1-1957, 78, D. e. ab. sublineata det. V. Laboissière, ZMUH [2006-02]; 1♂ diss., 2♂♂, 2♀♀, Taormina env. [ 37.85°N, 15.283333°E], 8-V-1942, Frey, NHMB [2003-20]; 1♂ diss., Toscani [ Toscana Province; specific locality not given], Tamarix gallica, Coll [ection] Chapuis, D. elongata det. V. Laboissière 1939, IRSNB [2006-09]; LEBANON: 1♀ diss., Beirut [ 33.87194°N, 35.50972°E], dunes south of, 10–15-V-1963, Kasy & Vartian, USNM [2003-20] [from series listed as D. elongata by Lopatin (1967)]; 4♂♂ diss., 1♀♀ diss., Beyrut [ Beirut], Syr, Krüper, NMHB [2003-12, 13, 17, 45, 48]; 1♀ diss., Beyrut [ Beirut], Syr, 20-IV-1936, Frey, NHMB [2003-38]; 2♂♂ diss., 1♀ diss., 1♀, Beyrut [ Beirut], Syrien, NHMB [2003-46, 2005-04], ZMHB [2003-09]; MACEDONIA: 2♀♀, Macedonien [specific locality not given], Emge., ZMAN; 1♂ diss., Mazedonien [specific locality not given], 4-VI-1987, J. Böhme, 533, D. elongata det. Erber 1985, ZMHB [2008-05]; 1♂ diss., 2♀♀, 1♂, Skopje env. [ 42.0°N, 21.4333°E], southern Yugoslavia, VI-1937, O. Kodyn, D. elongata det. Sterbud, NMPC [2004-52]; 1♂ diss., 1♀ diss., 5♂♂, 3♀♀, Strumica [ 41.4375°N, 22.64333°E], 19-V-1937, W. Liebmann, DEI [2003-20, 2005-01]; 1♀ diss., Vardar [ 42.00555°N, 21.32611°E; stream], D. elongata det. Burlini 1967, MSNM [2003-09]; MONTENEGRO: 1♂ diss., Sutomore [ 42.14277°N, 19.04666E], 22-V-1978, G.J. Slob, RBCN [2003-03]; PORTUGAL: 2♂♂ diss., [specific locality not given], Coll. Letzner, DEI [2003-10, 2006-01]; RUSSIA: 1♂ diss., Daghestan [Respublika Dagestan; specific locality not given], Leder. Reitter, 4358, Coll[ection] Kouril P5/46/62, NMPC [2005-40]; SPAIN: 2♀♀ diss., 1♀, Estepona [ 36.4333°N, - 5.1333°W], Malaga, 15-II-1982, H. Teunissen, RBCN [2003-13, 16]; SYRIA: 1♂ diss., [specific locality not given], D. e. var. carinata, MNMS [2004-04]; 1♀ diss, [specific localilty not given, D. elongata det. Le Moult, IRSNB [2006-11]; TURKEY: 1♂ diss., 1♀ diss., Adana [ 37.001667°N, 35.328889°E], D. elongata [ 1♂ diss., NHMB], HNHM [2005-02], NHMB [2003- 10]; 3♂♂ diss., 1♀ diss., 3♀♀, Adana, Asia Minor, Sterba, D. v. carinata det. TJS [ 1♀ undiss.], NMPC [ 2005-08–10, 12]; 1♀, Adana, Asia Minor, H. Rolle, ZMHB; 1♂ diss., Alanya, 10 km west [ 36.6°N, 31.89°E], 18-VII-1972, [on T. smyrnensis (Gerling and Kugler 1973) ], T18, TAUI [2003-04]; 1♀ diss., Anamur, 30 km west [ 36.0845°N, 32.6082°E], 17-VIII-1972, [on T. smyrnensis (Gerling and Kugler 1973) ], T17, TAUI [2003-09]; 1♀ diss., Ankara [ 39.927222°N, 32.864444°E], V-1937, Dr. Vasvari, D. elongata det. K. Lopatin 1961, HNHM [2004-08]; 1♂ diss., 2♀♀, Bergama, 5 km north [ 39.1553°N, 27.1629°E], 24-VIII- 1972, [on T. smyrnensis (Gerling and Kugler 1973) ], T38, TAUI [2003-06]; 3♂♂ diss., 1♀ diss., 1♂, 1♀, Bürücek [Burucek; 37.35°N, 34.83333°E], Toros [Mt.], Anat[olia], 29–31-VII-1947, Exp. N. Mus. CSR, NMPC [2004-08, 2005-07, 14, 15]; 1♀ diss., 1♂, Cordélio [Karsiyaka; 38.4614°N, 27.1119°E], D. elongata det. V. Laboissière 1939, IRSNB [2006-10]; 1♂ diss., 2♀♀, Ephesos [Selcuk; 37.9517°N, 27.3747°E], 18-V- 1992, V. Nemec, EGRC [2005-03]; 1♂ diss., 2♂♂, Ereckli [Erekli; 41.28944°N, 31.41805E], Asia Minor, Bodemeyer, 824 [ ZMHB], D. elongata [ ZMHB], ZMHB [2003-08], NMPC; 1♂ diss., 2♀♀ diss., Eski–Chehir [ Eskisehir; 39.77667°N, 30.52056°E], Asia Minor, Bodemeyer, NMPC [2004-29; 2005-29, 30]; 1♀ diss., Izmir, 10 km east [ 38.43°N, 27.19°E], 23-VIII-1972, [on T. smyrnensis (Gerling and Kugler 1973) ], T41, TAUI [2003-08]; 1♂ diss., Kash [Kas], 30 km west [ 36.27°N, 29.333333°E], 20-VIII-1972, [on T. smyrnensis (Gerling and Kugler 1973) ], T27, TAUI [2003-01]; 1♂ diss., Kassaba [Kasaba; 36.31111°N, 29.73472°E], Manisa Prov., Asia Minor, 1-VIII-1931, B.P. Uvarov, Brit. Mus. 1931-468, USNM [2004-01]; 1♂ diss., Malatya [ 38.353333°N, 38.311944°E], VI-1964, Seidenstücker, UH.G, DEI [2003-16]; 1♂ diss., 2♂♂, Ortakche [Ortakoy; 37.4167°N, 28.7167°E], east of Aydin on Menderes River, Asia Minor, 23-VII- 1931, B.P. Uvarov, British Museum 1931-468 [ ZIN], BMNH [2003-09], ZIN [ 1♂]; 2♀♀ diss., Selcuk, 10 km east [ 37.8871°N, 27.4106°E], 22-VIII-1972, [on T. smyrnensis (Gerling and Kugler 1973) ], T37, TAUI [2003- 05, 07]; 2♂♂ diss., Smyrna [ Izmir; 38.407222°N, 27.150278°E], 1955, Erwerb, Coll. Brancsik, NHMB [2003-09, 11]; 1♂ diss., 5♀♀, Tarsus [ 36.91778°N, 34.89167°E], Asia Minor, Sterba, NMPC [2004-47]; 1♂ diss., 1♀ diss., 1♀, Turgutli [Turgutlu; 38.5008°N, 27.7058°E], 65 km east Izmir, 23-VIII-1972, [on T. smyrnensis (Gerling and Kugler 1973) ], T40, TAUI [2003-02, 2004-01]; 1♂ diss., 1♀, Ushak, Abide [ Usak; 38.68°N, 29.40805°E], 3-VI-1989, Kuff & Szailles, RBCN [2003-01]; 1♀ diss., Yeniköy [multiple possible geocoordinates], Toros [Mt.], Anat[olia], 30-VIII-1947, Exp. N. Mus. CSR, NMPC [2005-06]; UKNOWN COUNTRY: 1♀ diss., Dobrudscha [Dobruja region; around border of Bulgaria and Romania by Black Sea ; specific locality not given; app. location: 43.6843°N, 28.5364°E], D. elongata , 126, Coll. Schultheiss, DEI [2005-06]; 1♀ diss., Tichakir [geocoordinates not locatable], Tal Noda, Asia Minor [possibly an archaeological site near the Aegean Sea coast of Turkey], J. Weise collection, ZMHB [2003-07]; UNITED STATES OF AMERICA (introduced): California: Yolo Co.: 1♂ diss., Cache Creek near Rumsey [ 38.8895°N, - 122.2333°W], 28-IX-2006, J. Herr, on T. parviflora , voucher [source: 3 km west Sfakaki (nr Stavramenos), Greece], GSWRL [2007-47]; Texas: Howard Co.: 1♂ diss., 4♂♂, 7♀♀, Higgins Ranch near Beals Creek, 32.251[069]°N, -101.386[506]°W, 8 km east of Big Spring, 18-VIII-2005, J.L. Tracy, on T. chinensis / T. canariensis [on regrowth following July defoliation by Diorhabda ], voucher [source: 3 km west Sfakaki (nr Stavramenos), Greece], GSWRL [2006-11]; 5♂♂ diss., Higgins Ranch near Beals Creek, 32.24966°N, - 101.38582°W, Big Spring, 1-IX-2006, J.L. Tracy, on Tamarix chinensis / T. canariensis, GSWRL [2008-01, 02, 03, 04, 05]; Potter Co.: 1♂ diss., 9♂♂, 5♀♀, Lake Meredith National Recreation Area, 35.52774°N, - 101.76322°W, 11-IV-2006, V. Carney, on T. ramosissima , voucher [source: Posidi Beach, Greece], GSWRL [2006-18].
Distribution. General. Diorhabda elongata was most frequently collected from Italy to Bulgaria and central Turkey, but it occurs sporadically elsewhere around the Mediterranean in Lebanon, Egypt, Algeria, Portugal and Spain, and near the Caspian Sea in southern Russia ( Dagestan Republic). Its native distribution is restricted to countries bordering on the Mediterranean Sea and the additional countries of Portugal, Bulgaria, Macedonia, and Russia (Map 2). Previous reports for the general distribution of D. elongata ( Weise 1924, Heyden et al. 1891, Winkler 1924 –1932, Ogloblin 1936, Wilcox 1971, Warchalowski 2003, Lopatin et al. 2004) are accurate only in their inclusion of the Mediterranean region to southern Russia. Further collections should provide specific localities of D. elongata along coastal areas of Portugal, France, Romania (Dobruja region), Syria, Algeria, and southern Russia ( Dagestan Republic). Additional collections could also reveal D. elongata as common in Slovenia and uncommon in Serbia, Georgia and the coastal areas of Azerbaijan, Tunisia, Libya and Morocco. The D. elongata group is not reported from Israel ( Lopatin et al. 2003).
Confirmed Records. We have dissected specimens of D. elongata from the following countries with previous literature records (Map 2): Portugal ( Corréa de Barros 1924), Spain (Hopkins and Carruth 1954, Torres Sala 1962, Petitpierre 1988), Italy (Reiche and Saulcy 1858, Weise 1893, Porta 1934, Zocchi 1971, Lundberg et al. 1987 b, Biondi et al. 1995), Greece ( Brullé 1832, Reich and Saulcy 1858, Regalin 1997), Bulgaria ( Tomov 1969, 1979, 1984; Warchalowski 1974; Gruev and Tomov 1986, 1998), Turkey (Reiche and Saulcy 1858; Ogloblin 1936; Kerville 1939; Gerling and Kugler 1973 [list on p. 20 should indicate Turkey (T), not Israel (blank) beside D. elongata ]; Aslan et al. 2000; Gök and Çilbiroǧlu 2003, 2005; Gök and Duran 2004), Russia ( Ogloblin 1936), Cyprus ( Georghiou 1977) , Syria (Reiche and Saulcy 1858, Ogloblin 1936), Lebanon (Reiche and Saulcy 1858 [as Syria], Ogloblin 1936 [as Palestine], Lopatin 1967 [as Lebanon]), Egypt (Reiche and Saulcy 1858, Weise 1925), Algeria (Reiche and Saulcy 1858, Peyerimhoff 1926), and the United States ( Texas, California; introduced; Map 7, see Potenial in Tamarisk Biological Control below for more details) ( Riley et al. 2003, DeLoach et al. in prep., Hudgeons et al. 2007a). We dissected D. elongata from a series of Beirut, Lebanon, identified as D. elongata by Lopatin (1967). A male D. elongata we dissected was collected in 1982 by Thure Palm from Fiumi di Pollina, Sicily, Italy, and it probably is associated with studies Palm co-authored reporting collections by himself and others ( Lundberg et al. 1987a) of D. elongata on T. gallica at Fiumi di Pollina ( Lundberg et al. 1987b).
New Records. We have dissected D. elongata from the following countries for which we find no previous specific reports in the literature: Croatia, Bosnia and Herzegovina, Montenegro, Macedonia (The Former Yugoslav Republic), and Albania.
Unconfirmed Records. We cannot confirm reports of D. elongata from Georgia (Reiche and Saulcy 1858, Lozovoi 1961) and Azerbaijan (Samedov and Mirzoeva 1985; Mirzoeva 1988, 2001). Because of the predominance of D. carinata in these areas, we consider these reports to primarily involve D. carinata (see D. carinata — Distribution below), and the presence of D. elongata needs further confirmation. However, we have dissected D. elongata from the Dagestan region of Russia along the Caspian Sea ( Fig. 19 View FIGURES 19–23 — Dagestan) and suspect that it is present in Georgia and Azerbaijan. Reports of D. elongata in areas east of the Caspian Sea in central Asia (east of 50°E) (e.g., Lopatin, 1977a, Medvedev and Voronova 1977b, Bieṅkowski 2004) should refer instead to D. carinata and D. carinulata .
France lies between two countries with confirmed host records, Italy and Spain (Map 2). Therefore, although we have not examined D. elongata from France, we still consider the general locality record of France as accurate (Reiche and Saulcy 1858, Bedel 1892, Laboissière 1934, Chatenet 2002). All locations from which specimens were dissected in Bulgaria (9), Greece (49; with the exception of a single male specimen of D. carinata considered as mislabeled from Attica, Greece), and Italy (15) were D. elongata and we consider all literature records of D. elongata from these countries as accurate (Map 2). We examined a specimen of D. elongata from Italy (ZMUH) with an identification label of D. e. ab. carinata by Laboissière and we consider Porta’s (1934) report of D. e. ab. carinata in Italy to be D. elongata . In Turkey, D. carinata is found as far west as 41.5°E and it is found near the south Turkish border in Halab, Syria at 37°E longitude (Map 3). Therefore, we are only confident in accepting literature records of D. elongata in Turkey that are in the area west of 35°E longitude, from which we dissected D. elongata from all 21 available locations (Map 2).
Below are 13 unconfirmed locality records of D. elongata that we consider as valid (Map 2):
BULGARIA: Blagoevgrad environs, near Struma River [ 42.01670°N, 23.08000°E], on Tamarix ( Tomov 1979) ; 1 specimen, Melnik, 4-V-1971, Stück ( Warchalowski 1974) [ D. elongata dissected from same location]; GREECE: Kaiafas [ 37.5167°N, 21.6000°E], Peloponnes [ Pelopónnisos Peninsula], west coast, 24- V-1995 (Serge Doguet, Fontenay–sous–Bois, France, pers. comm.); Plakias [ 35.200°N, 24.400°E], Crete, 13- 16-VI-1995; Rethimnon nom. [Rethymnon; 35.3647°N, 24.4714°E], Crete, 13-16-VI-1995; Sèlia [Sellia; 35.4000°N, 24.2330°E], Crete, 13-16-VI-1995, ( Regalin 1997; on Tamarix smyrnensis ); ITALY: Fiume de Pollina [ 38.01667°N, 14.6667°E], 15 km east Cefalu, Sicily, on Tamarix gallica [ D. elongata dissected from same location]; Fiume de Tusa [ 38.01667°N, 14.2667°E], 21 km east Cefalu, Sicily, on Tamarix gallica ( Lundberg et al. 1987b) ; Trieste environs [ 45.6395°N, 13.7876°E] ( Weise 1893); TURKEY: Asagigökdere village [ 37.59722°N, 30.82833°E], Isparta Province, 650 m [elev.], 12-V-2000 ( 2♂♂, 3♀♀), on Tamarix smyrnensis , 2-VII-2000 ( 3♂♂, 4♀♀), 10-X-2000 ( 1♂, 1♀), 4-V-2001 ( 3♂♂, 3♀♀) (Gök and Çilbiroǧlu 2003, Gök and Duran 2004); 4 specimens, Edirne [ 41.67440°N, 26.56080°E], 15 m [elev.], 6-V-1960 (Tomov and Gruev 1975); 10 specimens, Yozgat dintorm [ 39.8200°N, 34.8044°E], Yozgat [Ili], 1,300 m [elev.], 26- VI-1975, G. Osella ( Tomov 1984).
Because of the proximity to locations of D. carinata , the following two unconfirmed distribution records of D. elongata from east of 35°E in Turkey are uncertain (Map 2):
TURKEY: 1 specimen, Diyarbakir dintorm [ 37.9189°N, 40.2106°E], Diyarbakir [Ili], 5-VI-1971, G. Osella GoogleMaps ; 5 specimens, Bafra [ 41.56780°N, 35.9069°E], Samsum [Ili], 8-VII-1975, G. Osella ( Tomov 1984) GoogleMaps .
Discussion. Taxonomy. Diorhabda elongata (Brullé) was described in the genus Galeruca by Brullé (1832) from Morée (= Pelopónnisos peninsula), Greece. We dissected a total of 16 male and 11 female topotypes from 12 locations of the type locality, Pelopónnisos, Greece (Map 2). These males and females shared unique genitalic characters among the D. elongata group, comprising a distinct genitalic morphotype pair along with all other specimens examined from Greece and the surrounding countries of Italy, Albania, Macedonia and Bulgaria as well as western Turkey (see discussion below; Figs. 14 View FIGURES 14–18 , 19 View FIGURES 19–23 , 24 View FIGURES 24–28 , 29 View FIGURES 29–33 , 34 View FIGURES 34–38 , 39 View FIGURES 39–43 , 44 View FIGURES 44–47 ; Map 2). We are certain that this genitalic morphotype pair is a single species conspecific with D. elongata .
The endophallic sclerites of D. elongata bear several characters distinguishing them from those of the holotypes of D. carinata , D. carinulata , and D. meridionalis (as illustrated in Figs. 18–19 View FIGURES 14–18 View FIGURES 19–23 of Berti and Rapilly 1973) and other specimens of these species and D. sublineata examined in this revision (see Male- Genitalia above; Figs., 15–18, 20–23, 25–28, 30–33). Additional unique qualitative characters of the female genitalia distinguish D. elongata from other species in the D. elongata group (see Female- Genitalia above; Figs. 34–43 View FIGURES 34–38 View FIGURES 39–43 ). The distinctive genitalic characters of D. elongata are maintained in the same areas where D. sublineata , D. carinata and D. carinulata occur and near its abutting range boundary with D. meridionalis , and this is strong evidence for reproductive isolation between these species (see Biogeography below; Map 1, Table 8). Further evidence for reproductive isolation between D. elongata and several members of the D. elongata group is also found in previously discussed differences in component ratios of putative aggregation pheromones and reduced F2 hybrid egg viabilities.
Both the text and color habitus drawing accompanying Brullé’s (1832, Plate 44, Fig. 10 View FIGURES 10–13 ) original description lack any indication of striping or vittae on the elytra, but we find that D. elongata commonly has submarginal and substural vittae ( Fig. 1 View FIGURES 1–9 ). The elytral vittae in D. elongata are confined to the apical half of the elytra ( Fig. 1 View FIGURES 1–9 ), distinguishing them from some specimens of the other four species of the D. elongata group in which the elytral vittae, when present, may extend into the basal half of the elytra ( Figs. 5, 9 View FIGURES 1–9 ). We find that external characters previously used to distinguish D. elongata from the sibling species D. carinata and D. sublineata ( Weise 1883, 1890; Laboissière 1934, Porta 1934, Bechyné 1961, Warchalowski 2003) are too variable for species diagnosis (for further details, see Discussion — Taxonomy under D. carinata and D. sublineata ). We have dissected specimens of D. elongata that were misidentified by taxonomists using external diagnostic characters as D. e. ab. carinata in Italy, D. e. var. sublineata in Greece, and D. e. ab. sublineata in Italy (see Material examined). Gruev and Tomov (1986) provide a detailed description of external adult morphological characters for D. elongata but their given size range of 4.5–8 mm, is wider than the size range we find of 5.3–7.7 mm, and their range certainly includes that of sibling species that are smaller ( D. carinulata ) and larger ( D. carinata ) ( Table 2). The size range of 5.5–8 mm given elsewhere in southern Europe ( Porta 1934, Laboissière 1934) is closer to our observation regarding minimum size.
Mulsant (Mulsant and Wachanru 1852) described G. costalis Mulsant from Cilicia (along the coast of southwestern Turkey, towards Syria) from specimen(s) of unknown sex. The given body length is 5.6 mm, possibly making the type a male (see Table 2). The elytral vittae (as striae) are described beginning at a point 3/5 th the length of the elytra from the base (starting in the apical half) and ending at a point 6/7 th the length of the elytra from the base (ending near the apical tip). Confinement of the vittae to the apical half of the elytra agrees with our observations for D. elongata . We dissected D. elongata from all four locations available from the type locality of Cilicia (an area including Adana in southwest Turkey, Map 2). We dissected D. carinata and D. meridionalis from ca. 100 km southeast of Cilicia in Halab, Syria (Map 1). We follow Reiche and Saulcy (1858) in regarding G. costalis as a synonym of D. elongata . Other synonyms established by Reiche and Saulcy (1858) and Weise (1893) actually consist of the three valid species D. carinata , D. sublineata , and D. carinulata .
The number of spines on the endophallic sclerites and the shape of the elongate sclerite are fairly variable in D. elongata ( Fig. 19 View FIGURES 19–23 ) and males from certain series were often more similar to one another than those from other series. For example, a majority of males dissected from Promontorio de Gargano, Italy, and 60 km E. Thessaloniki, Greece had greater spination of both the elongate and palmate sclerites than males from Burucek, Turkey. However, the variability in spination is continuous (see Figs. 19 View FIGURES 19–23 and 29 View FIGURES 29–33 , Methoni, Greece), and we detected no geographic pattern. We have seen only a few series of specimens collected from identified species of Tamarix hosts and further investigations should be made into potential patterns in morphological variability associated with host species. Lohse (1989) found sympatric and phenologically differing Rumex and Polygonum host ecotypes of Galerucella aquatica that could be distinguished by the coloration of the last abdominal sternite but not by endophallic sclerites. Studies of genetic relationships among D. elongata populations collected from various Tamarix spp. in southern Europe are in progress (R. Carruthers, USDA/ ARS, Albany, CA, pers. comm.).
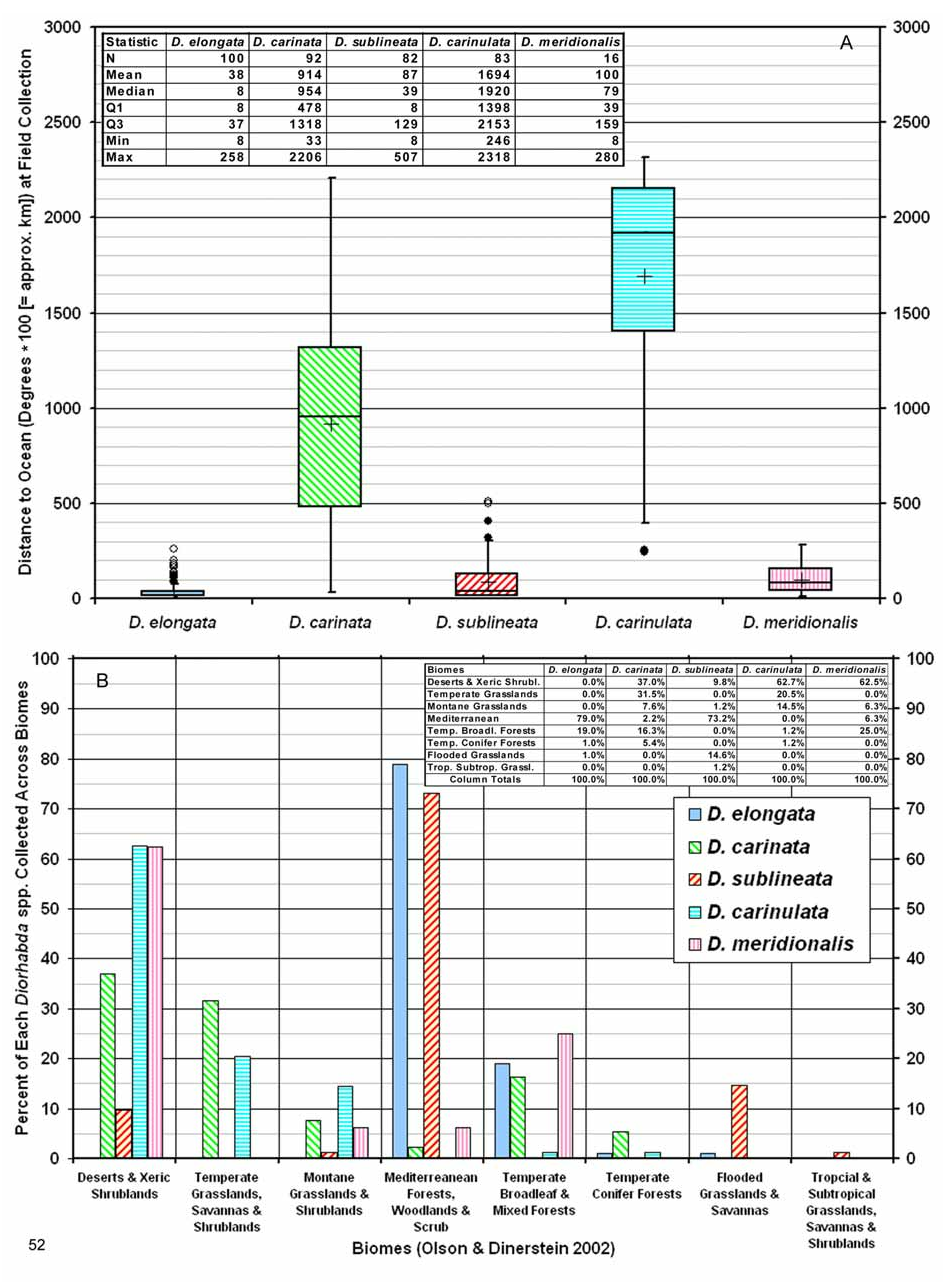
Common Name. The name “Mediterranean tamarisk beetle” refers to the great majority (ca. 80%; Fig. 52B View FIGURE 52 ) of collections of D. elongata being from the Mediterranean biome.
Biology. Host Plants. Diorhabda elongata has been collected from Tamarix smyrnensis Bunge , a close relative of T. ramosissima ( Baum 1978) , on Crete, Greece ( Regalin 1997) and in Isparta Province of southwest Turkey (Gök and Çilbiroǧlu 2003, 2005; Gök and Duran 2004) ( Table 1). Table 5 of Gerling and Kugler (1973) lists T. smyrnensis as the host species for seven locations in western Turkey associated with dissected TAUI specimens of D. elongata (see above Material Examined). Tamarix gallica is reported as a host in northern Sicily ( Lundberg et al. 1987b), and we dissected a specimen from the Toscana province of Italy with a host label of T. gallica . Diorhabda elongata is reported from Tamarix sp. in Cyprus ( Georghiou 1977) . New host records from collections on Peloponnisos, Greece by Javid Kashefi (USDA-ARS) are T. hampeana Boissier and Heldreich and T. parviflora . Tamarix parviflora is also a reported host from Cache Creek, California (DeLoach et al. in prep.; R. Carruthers, pers. comm.). Tamarix chinensis × T. canariensis / T. gallica is a new host record for populations of D. elongata established at Big Spring, Texas ( Table 1). Dalin et al. (in press) found that D. elongata from Crete preferred T. parviflora to a similar degree as T. ramosissima in multiple-choice field cage studies in California.
In Bulgaria, D. elongata severely defoliates Tamarix sp. trees (especially those in open areas) when it becomes numerous on the dry sandy terraces of the Struma River valley near Blagoevgrad ( Tomov 1979). Tamarix tetrandra Pallas , a close relative of T. parviflora ( Zieliṅski 1994), is common along the Struma River Valley and other parts of Bulgaria, Greece, and Turkey ( Baum 1978, Zieliṅski 1994), where it may serve as a host for D. elongata . Tamarix dalmatica Baum , the prevalent Tamarix species along the eastern coastlands of the Aegean Sea ( Baum 1978) , may serve as a host in Croatia, Montenegro and Albania. Reports of D. elongata in Central Asia from the leguminous shrubs Halimodendron ( Sinadsky 1960, Bieṅkowski 2004), Ammodendron ( Seitova 1974, Sinadsky 1968), and Alhagi ( Sinadsky 1968) (Fabaceae) should refer to Galerupipla sp. (see DeLoach et al. 2003b).
No-choice larval host suitability studies by Milbrath and DeLoach (2006a) confirm that D. elongata larvae from Crete can survive to adulthood only on plants of the order Tamaricales , including Tamarix (Tamaricaceae) and, to a lesser degree, on three North American Frankenia spp. (Frankeniaceae) : F.salina , F. johnstonii , and F.jamesii . Herr et al. (2006, in prep.) found larval survival on F. salina was not different than that on T. ramosissima . Multiple-choice adult oviposition studies in field cages revealed that the three North American Frankenia spp. provide little attraction for oviposition compared to Tamarix (Milbrath and DeLoach 2006a; Herr et al. 2006). Diorhabda elongata tended to oviposit less on T. aphylla compared to other invasive North American tamarisk, including T. ramosissima , T. chinensis , and T. canariensis / T. gallica in multiple choice field cage tests (Milbrath and DeLoach 2006 a, Herr et al. 2006). However, in other multiple-choice field cage tests among Tamarix spp. , oviposition by D. elongata on one accession of T. aphylla (Phoenix, Arizona) was not different than that on T. ramosissima × T. canariensis / gallica , T. canariensis / T. gallica and T. parviflora (Milbrath and DeLoach 2006b) . In a no-choice laboratory cage experiment, D. elongata accepted T. aphylla for oviposition to a significantly lesser extent than T. ramosissima but not T. parviflora ( Herr et al. 2006) . In this same no-choice experiment, the difference between oviposition by D. elongata on F. salina (inland variety) and T. ramosissima , T. parviflora , and T. aphylla was not significant ( Herr et al. 2006). Diorhabda elongata accepted T. aphylla for oviposition to the same degree they accepted T. ramosissima × T. chinensis in no-choice field cage studies (Milbrath and DeLoach 2006b). In open field testing with transplanted potted plants at Big Spring, Texas, D. elongata oviposited little on T. aphylla compared to T. ramosissima / T. chinensis (Moran et al. in press), and almost no oviposition occurred on F. salina ( Herr et al. 2006) . Tamarix aphylla is at moderate risk of damage by D. elongata in the field and it is difficult to predict to what degree D. elongata would damage T. aphylla , especially in the absence of other Tamarix spp. (Milbrath and DeLoach 2006b). Based on open-field testing in south Texas, Moran et al. (in press) concluded that D. elongata will likely have limited establishment and impact on T. aphylla . Frankenia is at very low risk of damage from D. elongata (Milbrath and DeLoach 2006a) . Risk of damage to both T. aphylla and Frankenia by D. elongata is probably much lower when these plants are not in the proximity of preferred Tamarix spp. (e.g., Blossey et al. 2001).
Ecology and Phenology. Tomov (1979) studied large populations of D. elongata from 1967–1971 along the Struma River near Blagoevgrad, Bulgaria. They report that the first eggs appear in early April and mating pairs are found from the middle of April until September. Larvae appear in early May until September, and the last adults are seen in October. Three population peaks of adults are seen near Blagoevgrad: late May and early June, late July and early August, and late September. Feeding by young larvae produces small holes in the lower epidermis and parenchyma of Tamarix leaves. Older larvae and adults eat entire leaves. Larvae pupate in loose cocoons in the soil. Late in the summer, many Tamarix shrubs, especially isolated ones, are completely defoliated along the Struma River by D. elongata .
Diorhabda elongata was noted in small numbers from May to October on T. smyrnensis in southwest Turkey (Gök and Çilbiroǧlu 2003). Adult collection dates from our examined material are from May to September in Croatia, April to October in Greece, and April to October in Italy. Our only specimens from Spain were collected on 15 February at Estepona on the southern coast.
Milbrath et al. (2007) found that D. elongata from Crete overwintering at Temple, Texas had ca. 80–95% survival from early November through the middle of March when tamarisk leaves began budding. Overwintered adults emerged in mid to late March and commenced ovipositing at the beginning of April giving rise to four generations and a partial fifth generation. Fourth generation adults emerging in early September oviposited little in September and ceased oviposition by October when they appeared to enter diapause.
In Big Spring, TX, five generations were also observed with overwintering adults generally emerging from late March and early April and fifth generation adults appearing from October to early November (DeLoach 2008, DeLoach et al. in prep.). On the Rio Grande from Candelaria to near Presidio, TX, large numbers of adults were actively defoliating trees into mid-November (A. Berezin, Sul Ross State University, Alpine, TX, pers. comm.). From Big Spring during 2005–2008, data was collected on the population density of D. elongata and associated rate of tamarisk defoliation on trees along sample transects throughout the growing season to develop models of wave dispersal using both a mathematical deterministic model and a statistical spatial regression model (J. Sanabria, pers. comm., DeLoach 2008, and DeLoach et al. in prep.).
Development and Reproduction. Egg mass size in the field in Bulgaria ranged from 10 to 19 eggs ( 26 in the laboratory) ( Tomov 1979). In laboratory studies at 28°C by Milbrath and DeLoach (2006b), D. elongata produced 16.0 ± 0.5 eggs per mass and fecundity averaged 219 ± 56.2 eggs with a population doubling time of 5.8 days on T. ramosissima × T. chinensis . On T. aphylla , fecundity was higher at 328.0 ± 60.4 but the population doubling time was longer at 6.6 days, which was partly due to both longer preoviposition and oviposition periods on T. aphylla . Milbrath et al. (2007) compared D. elongata from Crete with D. carinulata , D. carinata and D. sublineata at 28°C, and found D. elongata was similar to the other species with a development time of 21.0 days from egg to adult (with 78% survival), a fecundity of 281 eggs, and a population doubling time of 6.2 days.
Natural Enemies. Tomov (1974) reported the tachinid Erynniopsis antennata (Rondani) (= E. rondanii Townsend ) emerging from third instar larvae of D. elongata collected on 10 July in Bulgaria. E. antennata is also reported from southern France and Italy, and it also attacks the elm leaf beetle, Xanthogaleruca luteola (Müller) (Tschorsnig and Herting 1994) . Adult E. antennata oviposit on beetle larvae and tachinid adults emerge from late stage third instar larvae (prepupae) during the growing season and from overwintering beetles in the early spring (Dreistadt and Dahlsten 1990). In 1939, E. antennata was introduced from Europe and established in California for control of the exotic elm leaf beetle ( Flanders 1940). The parasitic fly has now spread throughout California ( Dahlsten et al. 1998), and is also reported from Oregon (USNM collection data). In California, parasitism rates of overwintering elm leaf beetle adults can reach 65%, but the fly is limited by a eulophid hyperparasite Baryscapus erynniae (Domenichini) (Dreistadt and Dahlsten 1990) . A protozoan mircorsporidian, Nosema sp. , was found in adult D. elongata originating from near Sfakaki, Crete, Greece (shipment EIWRU-2002-1002) (identified by J. Siegel) and Posidi Beach, Greece (shipment EIWRU- 2002-1009) (D. Bean, pers. comm.).
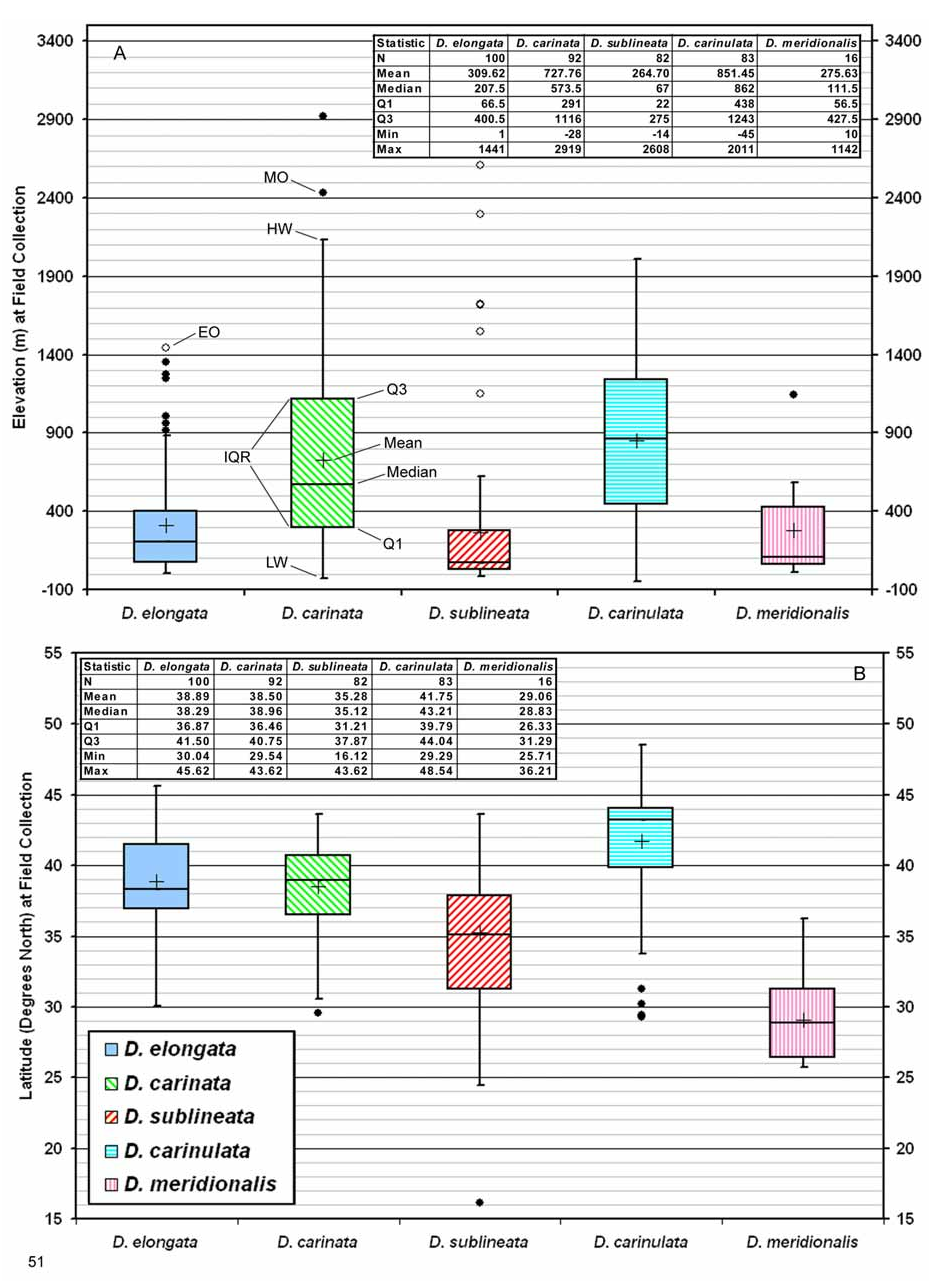
Biogeography. Comparative. Diorhabda elongata differs from other tamarisk beetles by the following combination of biogeographic characteristics: (1) strongly maritime and generally found within ca. 250 km from a sea coast, under 1,400 m elevation; (2) usually found in warm temperate Mediterranean woodlands or Temperate Broadleaf and Mixed forest biomes; and (3) latitudinal range of 30– 45°N and most common from 35– 43°N ( Table 7, Figs. 51–52 View FIGURE 51 View FIGURE 52 ). Diorhabda sublineata is marginally sympatric with D. elongata in Portugal, Spain and Egypt and both occur in the Mediterranean Forests, Woodlands and Scrub biome around the Mediterranean Sea (Map 1, Tables 8 and 9, Fig. 52B View FIGURE 52 ). However, D. elongata predominates in the northeastern Mediterranean in Europe and Asia while D. sublineata predominates in the western Mediterranean and all of North Africa, including more xeric biomes such as desert and flooded grasslands in which D. elongata is rare or absent. Diorhabda sublineata additionally differs from D. elongata in being common further south at 31– 35°N and ranging much further south to 16°N ( Figs. 51–52 View FIGURE 51 View FIGURE 52 ). Diorhabda are unreported along two ca. 600–700 km stretches of land around the Mediterranean where the main distributions of D. elongata and D. sublineata should interface in western Italy and Israel and Palestine, and no D. sublineata are reported from the main area of distribution of D. elongata (Map 1). If these D. elongata / D. sublineata interface zones actually lack Diorhabda populations, it may be worth investigating the potential existence of some form of localized competitive mutual exclusion between these species.
Diorhabda elongata is probably marginally sympatric with D. carinata in eastern Turkey, western Syria, and, possibly, southern Russia, Georgia and Azerbaijan. Diorhabda elongata is marginally sympatric with D. carinulata in southern Russia ( Dagestan) (Map 1; Table 8). Both D. carinata and D. carinulata are strongly continental and are mostly found in desert and grassland biomes from which D. elongata is not known. Diorhabda meridionalis is apparently parapatric with D. elongata in Syria and differs from D. elongata in being common both in deserts and further south at 26– 31°N ( Tables 7 and 8; Map 1; Figs. 51–52 View FIGURE 51 View FIGURE 52 ).
Descriptive. Diorhabda elongata is found in the west-central Palearctic realm (Maps 1). It is primarily collected from 35– 43°N in the northeastern Mediterranean from Italy to Bulgaria and western Turkey in two biomes: the Mediterranean Forests, Woodlands and Scrub and the Temperate Broadleaf and Mixed Forests (Map 2). Ecoregions (Olson and Dinerstein 2002) inhabited by D. elongata from 35– 43°N in the Mediterranean Forests, Woodlands and Scrub biome, based on frequency of collection, are the Ilyrian Deciduous Forests from Croatia to western Greece, the Crete Mediterranean Forests, and the Aegean and Western Turkey Sclerophyllous and Mixed Forests from coastal Greece to Turkey (Map 2). The latter ecoregion is the center of distribution for T. hampeana ( Boratyṅski et al. 1992, Zieliṅski 1994), a host plant of D. elongata . Tamarix hampeana occurs only rarely in the western Mediterranean area ( De Martis et al. 1986), similar to D. elongata (Map 2). The primary range of another host of D. elongata , T. parviflora , is also in the eastern Mediterranean (Map 2). In the Temperate Broadleaf and Mixed Forests biome, D. elongata inhabits the Balkan Mixed Forests from Macedonia to northwest Turkey. This area is a possible source for a putative northern interior lowland D. elongata climatype occurring from 41– 42°N and 200–500 m elev.
Potential in Tamarisk Biological Control. Summary. The Mediterranean tamarisk beetle is providing effective biological control of T. parviflora at Cache Creek, California and Tamarix ramosissima / T. chinensis near Big Spring and Pecos, Texas (Map 7). Based on its biogeographic characteristics, D. elongata is most suitable for Mediterranean biome of northern California ( Figs. 51–52 View FIGURE 51 View FIGURE 52 ; Map 13). D. elongata from Crete readily accepts the novel host T. chinensis × T. canariensis / T. gallica at Big Spring, Texas, and its absence in the deserts and grasslands of central Asia is probably due to poorer adaptation to the bioclimatic conditions of deserts and grasslands compared to other Diorhabda species rather than the lack of suitable Tamarix hosts. Diorhabda elongata will establish in west Texas grasslands and deserts around 31– 32°N, but our HSI model predicts other Diorhabda species are better biogeographically suited to deserts and grasslands of the southwestern U.S., including Diorhabda carinulata , D. carinata , and D. sublineata . More adapted Diorhabda species may eventually replace D. elongata where it establishes in desert and grassland habitats (Map 13).
Discussion. The Mediterranean tamarisk beetle attacks and defoliates Tamarix ( Tomov 1979) in Temperate Broadleaf and Mixed Forest biome at ca. 42°N in Bulgaria. Tamarix smyrnensis is a close relative of the invasive T. ramosissima in North America ( Baum 1978, Zieliṅski 1994), and it is attacked by D. elongata in Greece and Turkey. Diorhabda elongata attacks and damages both invasive T. parviflora and T. ramosissima / T. chinensis in North America. It has a moderate risk of damaging T. aphylla (Milbrath and DeLoach 2006b) and very low risk of damaging Frankenia (Milbrath and DeLoach 2006a) , and both these risks are probably much reduced at less proximity to preferred Tamarix spp. (e.g., Blossey et al. 2001).
The Mediterranean tamarisk beetle may be best adapted for areas from 35– 43°N in the maritime temperate warm Mediterranean Forests, Woodlands and Scrub biome and the Temperate Broadleaf and Mixed Forests biome (see Biogeography). Of these two biomes, only the Mediterranean Forests, Woodlands and Scrub biome is found in the western U.S. From 35– 43°N, the Mediterranean biome is represented by the California Interior Chaparral and Woodlands and the California Montane Chaparral and Woodlands ecoregions (Map 13). The Mediterranean species T. parviflora is the dominant invasive tamarisk in these ecoregions (Map 7). Diorhabda elongata from Crete has established well on T. parviflora near Cache Creek, California ( 39°N, Map 7), where it entirely defoliated over 200 hectares of tamarisk along an ca. 40 km reach in 2007, and over ca. 250 ha. (600 acres) along 50 km (including parts of nearby Bear Creek) in 2008 (DeLoach et al. in prep.; R. Carruthers, pers. comm.). This estimate of defoliated area is based upon an analysis of aerial photography which revealed ca. 396 ha of T. parviflora along a similar 40 km reach (ca. 10 ha of tamarisk per creek km) of Cache Creek in 2001 ( Ge et al. 2006). Tamarisk is estimated to occur at about half this density (ca. 5 ha. per km) over most of Cache and nearby Bear creeks where Diorhabda is well established (R. Carruthers, pers. comm.). Initial establishment at Cache Creek was slow, with very low populations persisting from 2004–2005, and only ca. 1.4 ha defoliated in 2006, and D. elongata has only weakly established and not established at some other northern California sites (John Herr, USDA/ARS, Albany, CA, pers. comm.). The possibility that establishment of D. elongata was slowed by predators or the need to adapt to a colder climate and to colder weather starting at longer daylengths earlier in the season should be investigated. At the Cache Creek site, potential parasitism of D. elongata by the previously discussed exotic parasitoid tachinid fly Erynniopsis antennata is being monitored (J. Herr, pers. comm.).
Diorhabda elongata from Crete is also well established on T. chinensis × T. canariensis / T. gallica (identified by J. Gaskin) in the Temperate Grasslands, Savannas and Shrublands biome (Western Short Grasslands ecoregion) near Big Spring, Texas ( 32°N, Map 7) ( Hudgeons et al. 2007a). During the first year of establishment at Big Spring in 2004, about four small trees were defoliated throughout the season, the first two trees being defoliated in July. In 2005, field populations increased and, by late September, over 200 tamarisk trees were totally defoliated, representing 0.17 ha of tamarisk canopy covered over a 0.66 ha area. By late October 2006, tamarisk over an ca. 7 ha area was defoliated ( Everitt et al. [2007] classified defoliation from remote sensing by color aerial photography), ca. 13 ha was defoliated in 2007, and over 40 ha was defoliated in 2008, including defoliation of an ca. 7 km stretch of Beals Creek with satellite populations defoliating scattered trees over a 19 km wide area around Big Spring. At each of two sites in the Trans-Pecos Chihuahuan Desert on the Pecos River near Pecos and Imperial, Texas (Map 7), ca. 1 ha of tamarisk trees were defoliated in 2007 during the second year post release (Mark Muegge, The Texas AgriLIFE Extension Service, Fort Stockton, TX, pers. comm.). By November 2008, defoliation by D. elongata along the Pecos River north of Pecos progressed to ca. 3.2 river km, but no defoliation was found near Imperial. Establishment was initially marginal or unsuccessful at other Texas AgriLIFE release sites in west Texas, but the use of ant baits in 2008 appears to be improving establishment at several of about 16 additional sites (A. Knutson, pers. comm.; see Map 7). Releases of populations of D. elongata from Crete were first made along the Rio Grande in Texas at several sites from Candelaria towards Presidio (Tyrus Fain, Rio Grande Institute, La Junta Project, Marathon, TX, pers. comm.) in the summer of 2007. Releases were made again in these areas in 2008 and in Big Bend National Park (Joe Sirotnak, Big Bend NP, TX, pers. comm.) and nearby Adams Ranch (M. Muegge, pers. comm.). Partial defoliation by D. elongata of several trees surrounding the release cages were noted at three sites between Presidio and Candelaria in the late summer and fall of 2008 (Mark Donet, USDA- NRCS, Alpine, TX, and Andrew Berezin, pers. comm.). In 2008, further releases were also made at various sites in west Texas, such as Iraan and Twin Buttes Reservoir near San Angelo (A. Knutson, pers. comm.), and at Holloman AFB, New Mexico (D. Thompson, pers. comm.).
In the Western Short Grasslands at Lake Meredith in north Texas ( 35°N, Map 7), D. elongata from Posidi Beach, Greece ( 40°N), overwintered in the open field after its release in 2005 and produced spotty damage to tamarisk, dispersing about 1 km, but it was present in only small numbers in 2008 (Erin Jones, Texas AgriLIFE Research, Amarillo, TX, pers. comm.). D. elongata from Crete failed to establish at Seymour, Texas in the Central and Southern Mixed Grasslands in 2004, but D. carinata from Qarshi, Uzbekistan was released there in 2008 and appeared to be establishing, defoliating over 0.2 ha during the first year of release (C. Randal, pers. comm.). In 2003, populations of D. elongata from Crete were released in the Chihuahuan Desert near Artesia, New Mexico ( 33°N, Map 7) and initially increased in 2004, but severely declined in early 2005 and could not be found through 2008.
Although the Mediterranean tamarisk beetle appears to be very promising for tamarisk biocontrol across the southern U.S. ( Milbrath et al. 2007), it has never been recorded in its native habitat from two biomes: Deserts and Xeric Shrublands and Temperate Grasslands, Savannas and Shrublands ( Table 9; Fig. 52B View FIGURE 52 ). We are not aware of a desert or grassland ecotype of D. elongata . During 2004–2007, D. elongata defoliated ca. 13.8 ha in the grassland biome at Big Spring compared to ca. 200 ha in the Mediterranean biome on Cache Creek. If the trend continues for more rapid defoliation in California, this could further support our biogeographic models showing that the Crete beetle is best suited to Mediterranean biomes. Other species of tamarisk beetles are common in desert and grassland biomes (Map 1, Table 9), and they may be better suited to much of the large area of these biomes invaded by tamarisk in North America as estimated by our relative Habitat Suitability Index Models (Map 13). Species distribution models incorporating climatic data are planned to better evaluate the potential suitability of D. elongata for climates across the western U.S. Making accurate predictions regarding potential North American ranges of any Diorhabda could be complicated if their field host preferences for Tamarix spp. vary significantly across their distributions.

| MNHN |
Museum National d'Histoire Naturelle |
| NMPC |
National Museum Prague |
| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
| NHMB |
Natural History Museum Bucharest |
| DEI |
Senckenberg Deutsches Entomologisches Institut |
| ZMAN |
Instituut voor Taxonomische Zoologie, Zoologisch Museum |
| NHRS |
Swedish Museum of Natural History, Entomology Collections |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| MZLU |
Lund University |
| CA |
Chicago Academy of Sciences |
| ZIN |
Russian Academy of Sciences, Zoological Institute, Zoological Museum |
| ZMUH |
Zoological Museum, University of Hanoi |
| CSR |
Caucasus State Nature Biosphere Reserve |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Diorhabda elongata ( Brullé, 1832 )
| Tracy, James L. & Robbins, Thomas O. 2009 |
Diorhabda elongata
| Porta, A. 1934: 317 |
Diorhabda elongata : Weise, 1883:316
| Hudgeons, J. & Knutson, A. E. & DeLoach, C. J. & Heinz, K. M. & McGinty, A. & Tracy, J. L. 2007: 157 |
| Carruthers, R. I. & Herr, J. C. & Knight, J. & DeLoach, C. J. 2006: 71 |
| Herr, J. C. & Carruthers, R. I. & DeLoach, C. J. 2006: 148 |
| Dudley, T. L. & Dalin, P. & Bean, D. W. 2006: 137 |
| Dudley, T. L. 2005: 13 |
| Dudley, T. L. 2005: 42 |
| Lopatin, I. K. & Aleksandrovich, O. R. & Konstantinov, A. S. 2004: 127 |
| DeLoach, C. J. & Carruthers, R. I. & Rodriguez-del & Bosque, L. A. 2003: 229 |
| Milbrath, L. R. & Herr, J. C. & Knutson, A. E. & Tracy, J. L. & Bean, D. W. & Rodriguez-del-Bosque, L. A. & Carruthers, R. I. & DeLoach, C. J. 2003: 225 |
| Chatenet, G. du 2002: 223 |
| Eberts, D. & White, L. & Broderick, S. 2001: 1 |
| Anonymous 2001: 52 |
| Aslan, I. & Warchalowski, A. & Ozbek, H. 2000: 30 |
| Campobasso, G. & Colonnelli, E. & Knutson, L. & Terragitti, G. & Cristofaro, M. 1999: 145 |
| Regalin, R. 1997: 69 |
| Biondi, M. & Daccordi, M. & Regalin, R. & Zampetti, M. F. 1995: 12 |
| Kovalev, O. V. 1995: 78 |
| Petitpierre, E. 1988: 93 |
| Lundberg, S. & Palm, T. & Trottestam, O. 1987: 126 |
| Georghiou, G. P. 1977: 47 |
| Warchalowski, A. 1974: 509 |
| Zocchi, R. 1971: 86 |
| Wilcox, J. A. 1971: 63 |
| Tomov, V. 1969: 181 |
| Lopatin, I. 1967: 441 |
| Jolivet, P. 1967: 331 |
| Torres Sala, J. de 1962: 327 |
| Kerville, H. G. 1939: 107 |
| Ogloblin, D. A. 1936: 79 |
| Laboissiere, V. 1934: 53 |
| Porta, A. 1934: 317 |
| Peyerimhoff 1926: 359 |
| Correa de Barros, J. M. 1924: 9 |
| Bedel, L. 1892: 158 |
| Heyden, L. V. & Reitter, E. & Weise, J. 1891: 375 |
| Weise, J. 1883: 316 |
Galeruca costalis
| Wilcox, J. A. 1971: 63 |
| Mulsant, E. & Wachanru, A. 1852: 176 |
Galeruca elongata Brullé, 1832:271
| Joannis, M. L. 1866: 83 |
| Brulle, A. 1832: 271 |