
Encyclia × tixtlensis C.E. León-Peralta, E. Salmerón-Barrera & E.A. Pérez-García, 2023
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.613.3.4 |
|
DOI |
https://doi.org/10.5281/zenodo.8356463 |
|
persistent identifier |
https://treatment.plazi.org/id/038C8031-2634-4D35-D1E2-FCB6FD0BFBF7 |
|
treatment provided by |
Plazi |
|
scientific name |
Encyclia × tixtlensis C.E. León-Peralta, E. Salmerón-Barrera & E.A. Pérez-García |
| status |
sp. nov. |
Encyclia × tixtlensis C.E. León-Peralta, E. Salmerón-Barrera & E.A. Pérez-García , nothosp. nov. ( Figure 1 View FIGURE 1 ).
Type:— MEXICO: Guerrero: Tixtla de Guerrero, in the transition zone between the tropical deciduous forest and the tropical oak forest, 1,400 m, October 27, 2019, C. León-Peralta & E. Salmerón-Barrera 50 (holotype: AMO!) .
Flowers similar to those of Encyclia meliosma but with proportionally larger lateral lobes, sepals and petals light to dark brown, lip brown at the base, purple on the apex and profusely magenta-streaked, colors that resemble E. atrorubens , but differs from it by the comical-pyriform pseudobulbs, the column bending at the basal third, and the fused lobes of the lip. This last feature also distinguishes it from E. hanburyi .
Description:— Litophytic herb, up to 31 cm tall excluding the inflorescence. Pseudobulbs grouped, conical-piriform, straight-very slightly arched, lightly compressed laterally, green, 6–8 × 3.5–5.0 cm. Leaves 2, dark green, sulcate on the adaxial surface, narrowly elliptical, obtuse, slightly reflexed, fleshy-coriaceous, 13–23 × 2.5–3.0 cm. Inflorescence up to 86 cm tall, a panicle with 1–3 branches, with 10–17 flowers opening simultaneously, peduncle elongated, smooth, up to 46 cm long. Floral bracts brown, 1–2 mm long. Flowers resupinate, 3.1–4.2 cm between the tips of the petals; sepals and petals pinkish brown to dark brown, veins slightly darker, light brown or yellowish at the base, lobes of the lip light to intense pink, with abundant magenta streaks, base of the lateral lobes brown, callus white, central base of the lip whitish-green; column white in general, dorsal surface with some yellowish brown spots, the longitudinal keel blushed with pink, ventral surface white, anther yellow. Sepals spatulate-oblanceolate, truncated at the base, apex obtuse, minutely apiculate, margins slightly wavy, 5-nerved; dorsal sepal 17–19 × 5.5–6.5 mm, lateral sepals somewhat falcate, slightly oriented backwards, with the basal third curved, 16–18 × 0.60–0.65 mm. Petals unguiculate, spatulate-oblanceolate, claw largely diffused longitudinally, apex obtuse-rounded, minutely apiculate, the margins slightly wavy, 15.5–18.0 × 6–7 mm, 1.0– 1.8 mm wide at the base. Lip trilobed, adnate to the column only at the base, the entire lip measuring 15.0–16.5 long and 15–18 mm wide between the apexes of the lateral lobes, the margins wavy-repandous; mid lobe 7 × 10 mm, transversely elliptical-flabellate, apex conduplicate, without a mucron, lateral lobes 9 × 4.5 mm, obliquely oblong-pandurate, falcate, adnate at the base to the medial lobe by 3 mm, apically rounded, apex erect and embracing the column; callus 5.7 × 2.6 mm, consisting of two thick keels forming a concave plate, obovate, smooth, sulcate, ending on a keel that is promptly thins out. Column appressed to the lip, dorsoventrally flattened, subtrigonate in cross-section, slightly arched upwards in the basal third, oblong-pandurate, slightly wider at the stigma, without auricles, with a longitudinal keel on the dorsal surface, the ventral surface sulcate, the entire column 7.5 × 4 mm; clinandrum 2 mm wide and 2 mm deep, 3-dented. Stigmatic cavity cordiform, sticky, 2 × 2 mm, the lateral lobes conspicuous; rostellum a transversal membrane, semi-elliptical, yellowish-cream, with viscarium on the ventral surface. Anther fleshy, subquadrate-cordiform, 4-locular, 1.8–2.0 × 1.5–2.0 mm. Pollinarium ca. 1.5 mm, with 4 obovoid pollinia, these very compressed laterally, yellow, 1 × 0.6 mm, joined in pairs to 1 mm long grainy caudicles. Ovary pedicellate, smooth, curved-sigmoid, 30–33 × 1–1.5 mm. Capsule elliptical, smooth, 5.6 × 1.9 cm (measurements of a developing capsule).
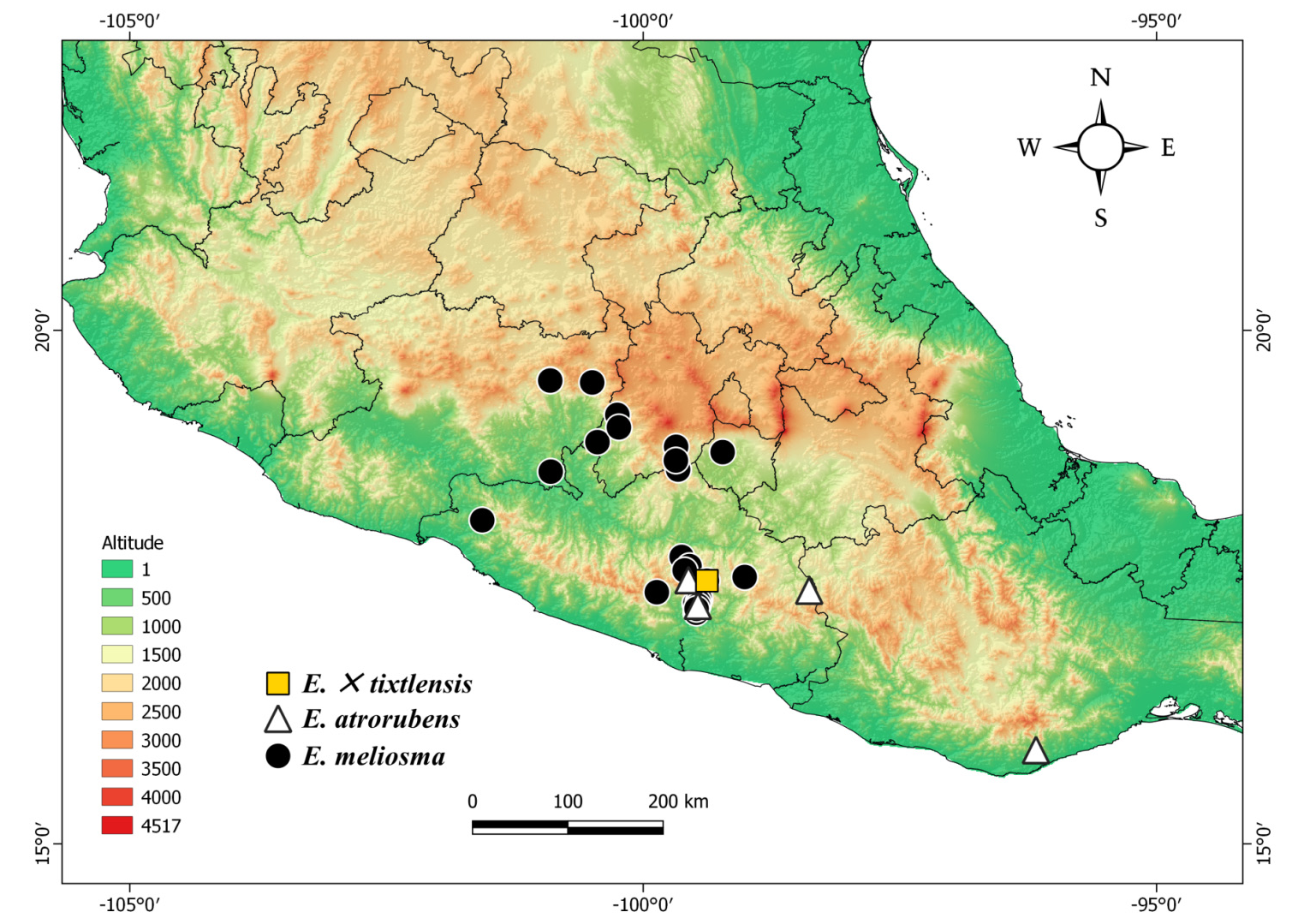
Distribution and ecology: —It is only known from the Tixtla de Guerrero region, Guerrero, México ( Figure 2 View FIGURE 2 ). Encyclia × tixtlensis grows between 1,400 -1,550 m as a lithophyte in the transition zone between the tropical deciduous forest and the tropical oak forest; it flowers from May to June in its natural habitat ( Figure 3a View FIGURE 3 ).
Etymology: —The specific epithet refers to the village of Tixtla de Guerrero, the place where the only specimens of the new taxon have been found. The name comes from nahuatl: “textli” = corn dough, and “tlan” = locality, which can be translated as “where there is corn dough”.
Additional specimens examined (paratypes): — MEXICO: Gerrero: Tixtla de Guerrero, aprox. 5 km al SO de Tixtla , ladera de exposición al sur, 1,550 m, 16 de junio de 1996, L. Cervantes 22 (MEXU!) .
Other records: — MEXICO: Tixtla de Guerrero, El Troncón , 1,550 m, abril de 2023, C. Jaimes sub C. León 300 (AMO, digital photo) .
Taxonomic discussion:—Plants of Encyclia × tixtlensis have conical-piriform, straight or slightly curved pseudobulbs, features that make it easy to differentiate it from E. atrorubens ( Figure 4 View FIGURE 4 ), whose pseudobulbs are rounded, green-purple, and from E. meliosma ( Figure 5 View FIGURE 5 ), which has laterally compressed piriform pseudobulbs. The flowers of E. × tixtlensis are similar to those of E. meliosma , but are clearly distinguished by the color, because the sepals and petals are light to dark brown, and the lip is brown on the base, purple on the apex and profusely magenta-streaked, in addition to the proportionally larger lateral lobes.
It could be thought to some extent that Encyclia × tixtlensis is part of the variation of E. meliosma . Soto-Arenas & Jiménez-Machorro (2002) pointed out that E. meliosma is not a species with any distinctive characters besides the fragrance; however, the specimens of the Tixtla de Guerrero region commonly lack a noticeable fragrance. Furthermore, E. meliosma belongs to a species complex that includes other very variable species, like E. rzedowskiana (Soto Arenas 2003: 565) and E. spatella .
Locally, E. meliosma is extremely variable, with lips with or without spots and stripes, and with apices round to markedly acuminate ( Figure 6 View FIGURE 6 ). Despite this, the vegetative morphology of the plants of E. × tixtlensis , as well as its geographic proximity to E. atrorubens lead us to think that this is rather a hybrid between E. atrorubens and E. meliosma .
Encyclia × tixtlensis shows a superficial resemblance to E. hanburyi , both in the morphology and in the color of the flowers, but the latter is easily distinguished by its wide, completely separated lateral lobes of the lip ( Figure 7 View FIGURE 7 ). Also, although E. hanburyi has been collected in Guerrero state (i.e. Alcozauca de Guerrero, Carnevali 6820 [CICY]), its localities are in eastern Guerrero, very far from the Tixtla region, making it a very unlikely parent of the new nothotaxon. Three Encyclia species have been found in the surroundings of Tixtla de Guerrero, namely E. meliosma , E. atrorubens and E. adenocaula (Lexarza 1825: 22) Schlechter (1918: 470) , which are found at different altitudinal ranges ( Figure 3 View FIGURE 3 ). Locally, E. meliosma is very abundant in the tropical dry forest under 1,400 m a.s.l. and may be considered to be exclusively lithophytic. On the other hand, E. atrorubens is found predominantly on oak trees over 1,400 m a.s.l. In the highest point of the area, E. adenocaula also grows as an epiphyte, with preference for oak trees. To this date, only two living plants of E. × tixtlensis have been located, both in the transition area between tropical dry forest and tropical oak forest, at approximately 100 m from the closest plants of E. atrorubens , but at less than 20 m away from plants of E. meliosma and E. adenocaula .
It is possible to discard Encyclia adenocaula as a possible parent of E. × tixtlensis since that species usually is very dominant in its hybrids, with its progeny inheriting attributes such as the long sepals and petals, and the warty inflorescences. A possible natural hybrid between E. adenocaula and E. meliosma was documented from the State of México by Szeszko (2011), but it’s notoriously different from the flowers of E. × tixtlensis in its star-shaped and pink flowers.Additionally, E. guadalupeae , an apparent hybrid described from the State of Nayarit between E. kennedyi and E. aenicta (the former being considered for some authors as a variety of E. adenocaula ( Hágsater & González-Tamayo 1975) , shows several distinctive characters of E. kennedyi ( Figure 8 View FIGURE 8 ).
A controversial aspect concerning the possible parents of Encyclia × tixtlensis is that E. meliosma flowers mainly in spring, while E. atrorubens flowers during fall. It is important to consider the high density of plants of E. meliosma that grow in the area, making it possible to find open flowers for a period of up to six months (E. Salmerón-Barrera, pers. obs.), facilitating the overlap with the first flowers of E. atrorubens in August (Leopardi, 2014), although it has been reported as blooming in July (Dressler & Pollard 1976). The mismatch between the flowering seasons between the possible parents might explain the rarity of E. × tixtlensis , since currently it is only known from the two living plants and an herbarium specimen (L. Cervantes 22 [MEXU]).
It has been suggested that Encyclia meliosma can also hybridize with other species; for example, Soto-Arenas (2002b) mentions the possible existence of hybrids between this species and E. huertae Soto Arenas & R. Jiménez (2002: 560) . Dressler and Pollard (1974) also suggested that a similar species, E. spatella ( Reichenbach 1856: 214) Schlechter (1924: 137) , hybridizes with E. hanburyi in the State of Guerrero. These authors based their hypothesis on a specimen collected on the road to Acapulco (UCBG[55479]). However, the flowers of that record show a yellowish color, and have lateral lobes of the lip united to the central lobe. Moreover, in this locality of Guerrero, E. hanburyi has not been found in the wild ( Leopardi-Verde, 2014). In our opinion, the suggested specimen corresponds to E. meliosma . Also, at the AMO herbarium there is a pressed flower identified as a hybrid between E. spatella and E. hanburyi (Coleman s.n.). The label does not include locality data, but it is mentioned that the plant was obtained as part of a package of E. cordigera ( Kunth 1815: 341) Dressler (1964: 247) . Encyclia hanburyi and E. cordigera are sympatric in southeast Mexico, where, to date, E. spatella has not been found. The flower of the specimen shows several characteristics of E. hanburyi , such as the long lobes of the lip separated from the central lobe, and in our opinion, it could correspond to that species.
Dressler & Pollard (1974) suggested a close relation between Encyclia atrorubens and E. diota ( Lindley 1843: 65) Schlechter (1918: 472) ; However, although both species belong to the Aptera clade of Encyclia , recent phylogenetic analyses based on DNA sequences have shown that E. atrorubens is more closely related to E. meliosma ( Leopardi-Verde et al. 2016, Carnevali et al. 2022). The Aptera clade represents a recent evolutionary radiation, with a high diversification rate coupled with a low relative extinction rate ( Leopardi-Verde 2014). Plants have been found in different regions whose morphology is considered intermediate between the species of this complex, some of which have been suggested to be natural hybrids (Soto-Arenas 2002). A developing fruit of E. × tixtlensis was found in the field, but it was not possible to determine the seed viability; nevertheless, this fact can contribute to the hypothesis that hybridization probably plays an important role in the diversification of the genus Encyclia .
Specimens examined of other Encyclia species: — Encyclia meliosma : Michoacán: M.A. Soto-Arenas 8755A, 8778, 8778, 8779, 8780 & E. Huerta (AMO), M.A. Soto-Arenas s.n. (AMO[26834]). Guerrero: J. Calónico-Soto 8818 (FCME), J. Calónico-Soto 13518 (FCME), M. Castañeda 728 & S. Ramos (MEXU), P:M: Catling M41 (AMO), R. Cruz-Durán 2248 (FCME), H. Flores 354 (FCME), E. Hágsater 1618 (AMO), E. Hágsater 3743 (AMO x7), E. Hágsater 3859 (AMO), M. Luna-Flores 863 (FCME, MEXU), A. González-Guinea 50 & A. Álvarez-Castro (FCME), M. Luna-Flores 867 (FCME), C. Figueroa 303, S. Ramos, M. Castañeda, J. Viccon & V. González (MEXU), H. Kruse 1045 (FCME), M.A. Soto-Arenas 10850 (AMO), D. Szeszko 295, 296, 299, 301, 302 & P. Szabó. Morelos: O. Nagel 3278 & J. González (AMO x2), E. Oestlund 1928 (AMO), J. Vázquez 3087 (MEXU). Estado de México: I. García-Cruz 437 (MEXU), G. Salazar 674 & M García-Rendón (AMO), R. Triay s.n. (AMO[4453]). Sin localidad específica: M.A. Soto-Arenas s.n. (AMO[26781]). Encyclia atrorubens : Guerrero: G. Carnevali 5915 (CICY), E. Hágsater 3745 (AMO), M.A. Soto-Arenas 10858 (AMO), Wrixon sub E. Hágsater 3733 (AMO, drawing). Oaxaca: J. Pascual 1939 (MEXU). Encyclia hanburyi : Guerrero: O. Nagel 7300 (AMES), O. Nagel 2284 & J. González (F, US), M.A. Soto-Arenas sub G. Carnevali 6820 (CICY). Oaxaca: A. García-Mendoza 10391, 10394, S. Franco, D. Sandoval & R. Avendaño-Rendón (MEXU), S. Maya 3379 (MEXU), E.A. Pérez-García 980 (MEXU), E.A. Pérez-García 1130 (MEXU), E.A. Pérez-García 1168, 1169, E. Durán & B. Reyes (MEXU), E.A. Pérez-García 2143 (MEXU), G.E. Pollard I-19o-31a (MEXU), S. Salas 1320 (MEXU).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |