
Monodelphis (Mygalodelphys) saci, Pavan & Mendes-Oliveira & Voss, 2017
|
publication ID |
https://doi.org/10.1206/3872.1 |
|
persistent identifier |
https://treatment.plazi.org/id/038C87AB-FF91-2163-2AA3-FE25FBE8FA53 |
|
treatment provided by |
Carolina |
|
scientific name |
Monodelphis (Mygalodelphys) saci |
| status |
sp. nov. |
Monodelphis (Mygalodelphys) saci , new species
Figures 4–8 View FIG View FIG View FIG View FIG View FIG
HOLOTYPE: The holotype consists of the skin, skull, postcranial skeleton, and preserved tissues of an adult male ( UFPA 1422 , original number JD 05 ) from Bom Jardim , Tapajós, Itaituba, Pará, Brazil ( 5.61° S, 57.12° W; fig. 2: locality 9), collected on 11 January 2013 by A.C. Mendes-Oliveira. GoogleMaps
PARATYPES: We refer 17 other examined specimens from the Brazilian state of Pará to Monodelphis saci , including: the skull, postcranial skeleton, and preserved tissues of a young adult animal of unknown sex ( MPEG 38947 View Materials [original number IAVRD 1135 ]) from Barragem da Pêra , Serra dos Carajás, Marabá ( 6.00° S, 50.21° W; fig. 2: locality 1) collected on 22 February 2005 by personnel from the Vale do Rio Doce Environmental Institute ; the skins, skulls, and postcranial skeletons of three adult males ( MCN-M 2301 , MN 73872 , MN 74003 [original numbers BM 72 , FMH 127, FMH 420]), one juvenile male ( MCN-M 1962 [original number ELP 007]), and one juvenile female ( MN 74002 [original number FMH 356]) from Floresta Nacional de Carajás , Parauapebas ( 6.05° S, 50.25 W; fig. 2: locality 2) 4; the skin, skull, and postcranial skeleton of an adult female ( MN 75511 [original number FMH 827]) from Floresta Nacional de Tapirapé-Aquiri , Marabá ( 5.80° S, 50.52° W; fig. 2: locality 3) collected on 24 February 2010 by D. Gettinger; the skin and preserved tissues of an adult male ( MPEG 40575 View Materials [original number CT 05]) from Rio das Tropas, Floresta Nacional do Crepori , Itaituba ( 6.52° S, 51.43° W; fig. 2: locality 4) collected on 29 April 2008 by S. Morato and L. Phov; a skin with skull in alcohol and preserved tissues of an adult female ( MPEG 42956 View Materials ) from Mina do Palito , Itaituba ( 6.31° S, 55.78° W; fig. 2: locality 5), collected on 21 March 2012 by A.O. Maciel and J.O. Gomes; the skin of an apparently juvenile male ( MPEG 41820 View Materials [original number CAC 308]) from Jardim do Ouro , Itaituba (ca. 6.27° S, 55.90° W; fig. 2: locality 6) collected on 4 April 2011 by C. Braga; the skin and skull of an adult male ( MZUSP 12207 View Materials [original number B-M-1]) from Uruá, Parque Nacional da Amazonia , Transamazon Km 65 ( 4.62° S, 56.25° W; fig. 2: locality 7) collected on 22 August 1978 by L.C. Branch; the skins, skulls, postcranial skeletons, and preserved tissues of two adult males ( UFPA 1394 , 1395 [original numbers JB 09, JB 10]) from Boca do Rato , Tapajós, Itaituba ( 5.23° S, 56.93° W; fig. 2: locality 8) collected on 19 January 2013 by A.C. Mendes-Oliveira; the skin, skull, and postcranial skeleton of an adult male ( UFPA 1260 [original number JMID 27 ]) and the skin and preserved tissues of an adult female GoogleMaps
4 Of these, MCN-M 2301 was collected on 18 December 2008 by R. Carvalho; MCN-M 1962 was collected on 16 January 2009 by E.L. Paschoalini; and MN 73872, 74002, and 74003 were collected respectively on 14 February 2009, 25 July 2009, and 25 February 2010, by D. Gettinger.
( UFPA 1738 [original number JD 33]) from Penedo, Tapajós, Itaituba (with two sets of coordinates [ 5.59° S, 57.12° W and 5.57° S, 56.13° W]; fig. 2: localities 10, 11) collected on 15 October 2012 and 17 June 2013 by A.C. Mendes-Oliveira; the skin, skull, and carcass in alcohol of an adult male ( UFSC 5210 [original number JC 3409]) from Marabá (specific locality unknown) collected on 25 January 2013 by G. Mette; and the skin of an apparently juvenile female ( MPEG 42601 View Materials [original number L079]) from São Félix do Xingu , (specific locality unknown) collected on 27 October 2011 by L.H. Diniz and T. R. Diniz GoogleMaps .
OTHER SPECIMENS: Four additional Brazilian specimens, apparently representing this species, have been reported in the literature, but we were not able to examine them for the present study. Among these, Semedo et al. (2011) reported one specimen ( UFMT 1355 ) from Fazenda São Nicolau , Mato Grosso ( 9.85° S, 58.22° W; fig. 2: locality 12), collected in December 2009 GoogleMaps . Additionally, Pavan et al. (2014) analyzed DNA sequences from one specimen ( UNIR M451 ) from Madeflona, Floresta Nacional do Jamari , Rondônia ( 9.14° S, 63.00° W; fig. 2: locality 13), collected in February 2012 GoogleMaps by R. Mendonça and M . R. Messias. Lastly, Abreu-Júnior et al. (2016) reported two specimens ( LMUSP 279 , 280 ) from Estação Ecológica do Rio Acre, Acre ( 10.75° S, 70.52° W; fig. 2: locality 14) collected in April 2015 and February 2016 GoogleMaps .
ASSOCIATED SEQUENCE DATA: DNA sequences from one mitochondrial gene (cytochrome b), two autosomal exons (IRBP exon 1, BRCA1 exon 11), one autosomal intron (SLC38 intron 7), and one X-linked intron (OGT intron 14) obtained from MPEG 40575 and MPEG 42956 were included in the phylogenetic analyses of Pavan et al. (2014) and are deposited in GenBank with accession numbers KM071402 View Materials , KM071124 View Materials , KM071035 View Materials , KM071316 View Materials , and KM071221 View Materials (corresponding to sequences obtained from MPEG 40575), and KM071403 View Materials , KM071123 View Materials , KM071034 View Materials , KM071315 View Materials , and KM071220 View Materials (corresponding to sequences obtained from MPEG 42956). DNA sequences from cytochrome b, BRCA1 exon 11, and OGT intron 14 loci of MPEG 38942 were also included in the phylogenetic analyses of Pavan et al. (2014) and are deposited in GenBank with accession numbers KM071404 View Materials , KM071036 View Materials , and KM071317 View Materials .
DISTRIBUTION: Monodelphis saci is currently known from at least 14 localities scattered along the south bank of the Amazon in the Brazilian states of Pará, Mato Grosso, Rondônia, and Acre (fig. 2).
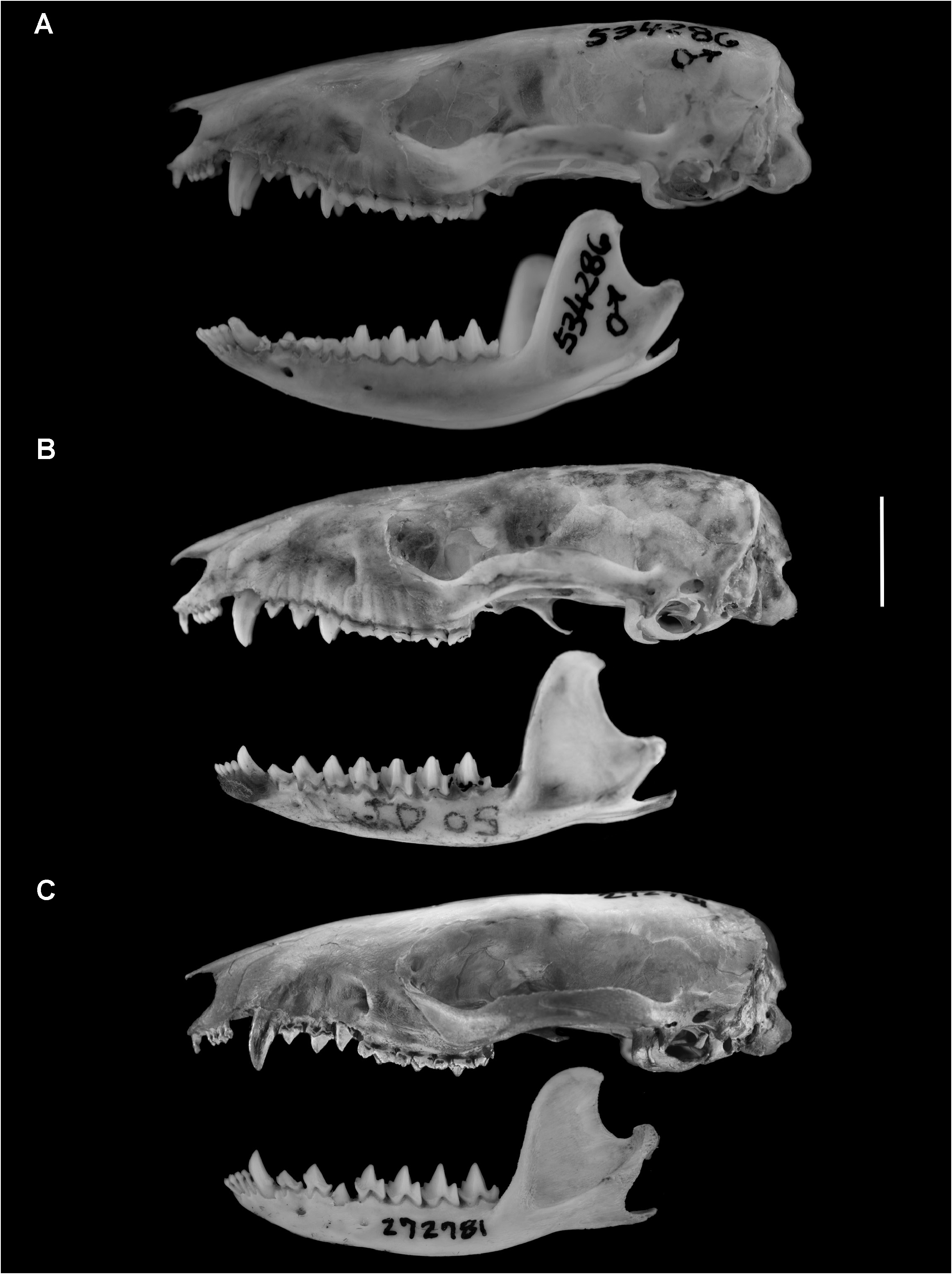
DESCRIPTION: Dorsal pelage superficially brownish from behind ears to rump (varying from near Clove Brown to Olive Brown, slightly darker on the rump than over the shoulders), without distinct stripes or other sharp pigmental discontinuities, but head conspicuously reddish (near Sepia); middorsal fur 4–5 mm long and grayish basally. Ventral pelage gray-based and usually brownish on most of chin, throat, groin, and flanks, but abruptly self-whitish midventrally on chest and abdomen. 5 Pinnae macroscopically naked, but densely covered internally and externally with short, brown hairs. Mystacial and supraorbital vibrissae mostly dark, with longest mystacial hairs extending behind eyes when laid flat against cheek but not reaching pinnae. There are two supraorbital vibrissae on each side (but only one is present on the right side on MPEG 42601 and UFPA 1422). Genal vibrissae lighter colored than the remaining facial vibrissae, usually reaching the pinnae but rarely extending behind their posterior border. Gular gland (indicated by an area of yellowish fur on the throat) present in all mature males and, apparently, in one out of three mature females examined. Hands and feet covered dorsally with short, mostly light-brown hairs. Ungual tufts very short (restricted to the bases of the manual claws and reaching only the middle of the pedal claws). Thenar and first interdigital pad of pes separate (not fused). Hypothenar pad of pes present (e.g., on MCN-M 2301) or absent (e.g., on MPEG 42956). Scrotal skin light brownish to cream, covered with pale (near Tilleul-Buff) hairs. Mammary formula apparently 3–0–3 = 6 (MPEG 42956), all abdominal-inguinal. Tail about half as long as combined length of head and body (mean LT/HBL = 0.51 in 8 males, 0.46 in three females; tables 2, 3), dark brown dorsally but mottled ventrally with pale blotches in mature specimens. Body pelage extends onto tail farther ventrally than dorsally. Caudal scales (discernible mostly on the distal portion of the tail) arranged in predominantly spiral series.
Skull unremarkable in general aspect (not conspicuously flattened; rostrum neither greatly elongated nor unusually short). Infraorbital foramen dorsal to anterior root of M1; anterior margin of lacrimal dorsal to M2 or to M1/M2 commissure; two lacrimal foramina usually present on anterior orbital margin. Interorbital region hourglass shaped, with rounded supraorbital margins, and lacking distinct processes, but supraorbital surface of frontals with
5 The ventral pelage of MPEG 40575, MZUSP 12207, and UFSC 5210 is dirty cream in general aspect, such that this marking is absent or indistinct.
V-shaped temporal scars in large males (e.g., UFPA 1395, UFSC 5210, MZUSP 12207); sagittal crest absent even in largest adult males examined; nuchal (occipital) crest present, restricted to dorsolateral margin of the skull (interrupted middorsally); no parietal-mastoid contact on posterior braincase (interparietal extends laterally to contact squamosal, except on left side of MZUSP 12207); zygomatic arches laterally expanded, with anterior connection to skull marked by shallow but distinct inflection in all examined mature specimens; frontal process of jugal absent or indistinct.
Incisive foramina extending from level of third upper incisors to between upper canines (never behind C1s); maxillopalatine fenestrae extending from M1 to posterior margin of M2 or middle of M3; anterior margin of maxillopalatine fenestrae usually posterior to infraorbital foramen in mature specimens (but ventral to the foramen on UFPA 1394); palatine fenestrae present (e.g., in MPEG 38947; MN 73872, 75511) or absent (e.g., MZUSP 12207; UFPA 1260, 1395); posterolateral palatal foramina small (not extending anteriorly between M4s) and complete (with an entire posterior bony margin); basisphenoid concealed by alisphenoid in lateral view (not exposed through sphenorbital fissure in posteromedial orbit); infratemporal crest of alisphenoid usually distinct; tympanic wing of alisphenoid small, uninflated, and widely separated from rostral tympanic process of petrosal; rostral tympanic process narrow, triangular, and widely separated from caudal tympanic process (fenestra cochleae exposed); anterior process of malleus exposed between ectotympanic annulus and tympanic wing of alisphenoid; extracranial course of mandibular nerve not enclosed by bone (secondary foramen ovale absent); stapes columelliform and imperforate; subsquamosal foramen large (substantially wider than the squamosal strut which separates this opening from the postglenoid foramen); parocciptal process elongated, dorsoventrally directed.
Crowns of I2–I5 increasing in width from front to back (such that the crown of I5 is distinctly wider than the crown of I2); upper canines usually simple, without distinct accessory cusps (but tiny accessory cusps are sometimes present in juveniles and young adults; e.g., in MN 74002, 74003, 75511); postcanine diastemata very small or absent in upper dentition; P2 and P3 usually subequal in width, or P3 slightly wider than P2 (e.g., on UFPA 1260 and 1422).
Lower dentition also without distinct postcanine diastemata; p2 and p3 subequal in height; anterior cingulid of lower molars narrow (on m2 and m3 these shelves are unambiguously lingual to the hypoconids of m1 and m2, respectively); lower milk premolar (dp3) narrow, with an incomplete (bicuspid) trigonid and a minimally differentiated anterior cingulid (MN 74002); entoconids of m1–m3 small or indistinct.
COMPARISONS: Monodelphis saci shares the diagnostic external and craniodental traits of the subgenus Mygalodelphys and therefore differs from species in other subgenera by having body fur that extends onto the tail farther ventrally than dorsally, lacking a distinct frontal process of the jugal, lacking parietal-mastoid contact on the lateral braincase, having a small sphenorbital fissure through which the basisphenoid is not exposed laterally, having narrow anterior cingulids on m2 and m3, and having indistinct entoconids on m1–m3 ( Pavan and Voss, 2016). Additionally, M. saci can be distinguished externally from sympatric congeners by its much smaller size and by its lack of conspicuously patterned dorsal pelage: whereas M. saci is uniformly brownish with reddish highlights only on the head, M. glirina ( Wagner, 1842) and M. touan ( Shaw, 1800) have reddish flanks, and M. emiliae ( Thomas, 1912) has a grayish midbody contrasting with its reddish head and rump.
Monodelphis saci can be distinguished from other species of the subgenus Mygalodelphys by a distinctive range of morphometric variation (tables 2, 3) 6 as well as qualitative external and craniodental traits. Among the latter, no other species of Mygalodelphys has a distinctly reddish head, nor does any other congener have pale mottling on the underside of the base of the tail. Other relevant comparisons with species of Mygalodelphys are summarized below.
Monodelphis saci can be readily distinguished from M. handleyi Solari, 2007 , and M. ronaldi Solari, 2004 , by its much smaller size (most external and all craniodental dimensions of
6 For comparative measurements of M. handleyi , M. ronaldi , M. reigi , M. osgoodi View in CoL , M. pinocchio , and M. kunsi View in CoL see Pavan (2015: table 1).
M. saci exhibit nonoverlapping variation with our samples of M. handleyi and M. ronaldi ). In addition, M. saci lacks several traits exhibited by M. handleyi and M. ronaldi , including an extremely flattened skull, a prominent sagittal crest, and a nuchal crest that is continuous across the dorsal midline of the occiput.
On average Monodelphis saci is smaller than M. reigi Lew and Peréz-Hernández, 2004 , in all measured external and craniodental dimensions, and is consistently smaller in four dimensions (HBL, TL, CBL, PL) that exhibit nonoverlapping variation in these species. In addition to the aforementioned differences in pelage color ( M. reigi is uniformly dark brown), M. saci lacks the conspicuously elongated rostrum and the distinct postcanine diastemata of M. reigi .
By comparison with Monodelphis osgoodi ( Doutt, 1938) and M. pinocchio Pavan, 2015 , M. saci can be distinguished, among other features, by a much broader and less elongated rostrum (measurements of PB exhibit nonoverlapping variation in pairwise comparisons of M. saci with either species), by the absence of distinct postcanine diastemata in the upper and lower dentition, and by shorter maxillopalatine fenestrae that extend anteriorly only to M1 (maxillopalatine fenestrae extend anteriorly to P3 or to the P3/M1 commissure in M. osgoodi and M. pinocchio ).
Monodelphis saci is substantially larger than M. kunsi Pine, 1975 View in CoL , in all measured external and craniodental dimentions (e.g., males of these species exhibit no overlap in HBL, CBL, PL, PB, and LM). In addition to its reddish head and pale-mottled caudal undersurface, M. saci also differs from M. kunsi View in CoL by its longer dorsal pelage ( 4–5 mm versus 3 mm in M. kunsi View in CoL ), tail scales in predominantly spiral series (annular series predominate in M. kunsi View in CoL ), a more elongated rostrum (best appreciated in side-by-side cranial comparisons), distinct lateral inflections where the zygomatic arches join the rostrum (absent in M. kunsi View in CoL ), a reduced facial exposure of the lacrimal (not extending anteriorly beyond the M1/M2 commissure, versus dorsal to M1 or extending to the P3/M1 commissure in M. kunsi View in CoL ), and upper incisor crowns that increase in width from I2 to I5 (versus I2–I5 crowns subequal in M. kunsi View in CoL ).
Monodelphis saci is similar to M. adusta View in CoL and M. peruviana ( Osgood, 1913) in most external and craniodental dimensions, but differs in qualitative traits. In addition to its reddish head and pale-mottled caudal undersurface, M. saci exhibits distinct lateral inflections at the anterior roots of the zygomatic arches (absent in M. adusta View in CoL and M. peruviana ) and shorter maxillopalatine fenestrae, with the anterior margin of these openings usually posterior to the infraorbital foramen (the anterior margin of the maxillopalatine fenestrae are usually anterior to the infraorbital foramen in M. adusta View in CoL and M. peruviana ). Additionally, we observed V-shaped temporal scars on the supraorbital surface of the frontals in most large male specimens of M. saci but not in any examined material of M. adusta View in CoL or M. peruviana .
HABITATS AND SYMPATRY: At Bom Jardim, Penedo, and Boca do Rato, Monodelphis saci was found in both primary and disturbed forest (logged areas and secondary vegetation) on both banks of the Rio Tapajós. Capture sites at these localities included terra firme forest characterized by a relatively low canopy ( 10–15 m) with numerous lianas (fig. 3A–B), as well as seasonally flooded riparian forest characterized by clayey soil, abundant epiphytes, palm trees, and herbaceous vegetation (fig. 3C).
Monodelphis saci probably occurs sympatrically with M. emiliae and M. glirina throughout its known geographic range, and it occurs sympatrically with M. touan between the Xingu and Tocantins rivers. Sympatry among M. saci , M. glirina , and M. touan has previously been reported from Floresta Nacional de Carajás ( Martins et al., 2012), whereas sympatry among M. saci , M. glirina , and M. emiliae was recently reported from the upper Rio Acre (Abreu- Júnior et al., 2016). Monodelphis saci and M. glirina occur syntopically along the middle Tapajós (where both species were caught in three lines of pitfall traps by the second author), and it occurs syntopically with M. emiliae at Mina do Palito (where the two species were caught in the same line of pitfall traps by A.O. Maciel and J.O. Gomes; personal commun.).
REMARKS: Several specimens deposited at the Museu Nacional (MN 73872, 74002, 74003) that we refer to Monodelphis saci were previously identified as M. kunsi by Gettinger et al. (2011).
ETYMOLOGY: The specific epithet is a noun in apposition and refers to the Brazilian folkloric character Saci, a one-legged gnome with a red cap. Saci is allegedly derived from the Yaci Yaterê of Tupi-Guarani mythology, to which elements of African and European folklore have been added over the last several centuries ( Cascudo, 1947).
OTHER SPECIMENS EXAMINED: Monodelphis adusta ( N = 24)— COLOMBIA, Boyacá, Guaicaramo ( AMNH 75232 About AMNH ) ; Cundinamarca, Paime ( MNHN 1929-672 About MNHN ) , Susumuco ( BMNH 21.5.2.6), “W. Cundinamarca ” ( BMNH 97.7 .2.1 [ holotype of adusta ]) ; Magdalena, Colonia Agricola de Caracolicito ( USNM 280894 About USNM ) ; Meta, Villavicencio ( AMNH 136158 About AMNH , 139227 About AMNH , 202650 About AMNH ) , 7 km NE Villavicencio ( AMNH 207765 About AMNH ) ; Tolima, Mariquita ( BMNH 12.4 .2.10, 12.9.8.3) . ECUADOR, Napo, Lumbaqui ( USNM 534286 About USNM ) , San Jose Abajo ( AMNH 68136 About AMNH ) ; Pastaza, Mera ( AMNH 67274 About AMNH ) , Safari Hostería Park ( TTU 84865, 84899 View Materials ) ; Tunguragua, Palmera ( AMNH 67275 About AMNH ) ; Zamora-Chinchipe, Zamora ( AMNH 47189 About AMNH ) . PANAMA, Darien, Cana ( USNM 179609 About USNM [ holotype of melanops]), Guayabo ( ANSP 19676 About ANSP ) , Tacarcuna Village Camp ( USNM 309263 About USNM ) . PERU, Loreto, 1.5 km N Teniente Lopez ( KU 157978) , Estación Biológica Allpahuayo ( TTU 98923, 101164 View Materials ) .
Monodelphis handleyi ( N = 6)— PERU, Loreto, Jenaro Herrerra (AMNH 276698, 276704, 276709; MUSM 23808–23810).
Monodelphis kunsi ( N = 36)— ARGENTINA, Salta, Finca Falcón (MACN-Ma 23783, 23784) . BOLIVIA, El Beni, La Granja ( USNM 461348 About USNM [ holotype of kunsi ]) ; Santa Cruz, El Refugio ( USNM LHE 1692 ) ; Tarija, Río Lipeo ( ANSP 18191 About ANSP ) , Tapecua ( AMNH 263968 About AMNH ) . BRA- ZIL, Distrito Federal, Fazenda Agua Limpa ( OMNH 22265–22266 View Materials ) ; Goiás, Silvânia ( MN A 191 ) ; Mato Grosso , Fazenda Tanguro ( MPEG 43016–43017 View Materials ) , Reserva do Patrimonio Particular Natural SESC Pantanal ( MN 64323 , 64411 , 64424 ) ; Mato Grosso do Sul , margem direita Córrego Urucum ( UFMG LFS 202 ) , Mineração Rio Tinto ( UFMG RM 318–319 , 324 View Materials , 340 View Materials ) ; Minas Gerais, Aiuruoca ( MCN-M 1570 , 1607 ) , Betim ( MCN-M 1470 , 1531 ) , Divinópolis ( MCN-M 1256 ) , Palmital ( MCN-M 2964 ) , Parque Nacional da Serra do Cipó ( MZUSP MTR 20361, MCN-M 1465 ) , Pompéu ( MCN-M 1420 , 1421 ) , Rio Manso ( MCN-M 2273 ) , Serra do Salitre ( MCN-M 2304 ) , Unidade Ambiental Jacob ( UFMG 1965 View Materials ) ; São Paulo , Nadiu Figueredo ( MN 46571 ) ; Tocantins, Peixe ( MZUSP 35059 View Materials ) . PARAGUAY, Canendeyú, Reserva de Biosfera del Bosque Mbaracayú ( TTU 116526 View Materials ) ; Presidente Hayes, Trans-Chaco Highway, Km 412 ( FMNH 164094 About FMNH ) . Locality unknown ( MN 73381 ) .
Monodelphis osgoodi ( N = 3)— BOLIVIA, Cochabamba, 4.4 km N Tablas Monte ( AMNH 264922 About AMNH ) , Incachaca ( CM 5242 [ holotype of osgoodi ]) . PERU, Cusco, Ocobamba Valley ( USNM 194379 About USNM ) .
Monodelphis peruviana ( N = 13)— BOLIVIA, La Paz, Serranía Bella Vista (AMNH 264562). PERU, Cusco, 2 km SW Tangoshiari (USNM 588019), 3 km E Amaybamba (MVZ 173928), Cashiriari (USNM 582782), Pillahuata (FMNH 172032), Suecia (FMNH 189811); Huánuco, Hacienda San Antonio (USNM 259433); Junín, Cordillera Vilcabamba (USNM 582110); Loreto, Nuevo San Juan (AMNH 272695, 272781; MUSM 13297, 15318); Madre de Dios, Reserva Cusco Amazónico (MUSM 7157).
Monodelphis pinocchio ( N = 7)— BRAZIL, Espírito Santo, Serra do Caparaó ( MZUSP MTR 15815) ; Rio de Janeiro, Alto do Itatiaia ( AMNH 61547 About AMNH ) , Serra de Macaé ( MZUSP 2793 View Materials ) ; São Paulo, Reserva Florestal do Morro Grande ( MN 78651 , 78680 [ holotype of pinocchio ]), Riacho Grande ( MZUSP 30740 View Materials ) . Locality unknown ( MZUSP 33878 View Materials ) .
Monodelphis reigi ( N = 2)— GUYANA, Potaro-Siparuni, Mount Ayanganna ( ROM 114699 About ROM , 114864 About ROM ) .
Monodelphis ronaldi ( N = 1)— PERU, Madre de Dios, Pakitza ( MUSM 17027 ) .




| R |
Departamento de Geologia, Universidad de Chile |
| MN |
Museu Nacional, Universidade Federal do Rio de Janeiro |
| T |
Tavera, Department of Geology and Geophysics |
| TTU |
Texas Tech University, Museum |
| KU |
Biodiversity Institute, University of Kansas |
| USNM |
Smithsonian Institution, National Museum of Natural History |
| ZIL |
Academy of Sciences, Zoological Institute |
| UFMG |
Universidade Federal de Minas Gerais |
| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Monodelphis (Mygalodelphys) saci
| Pavan, Silvia E., Mendes-Oliveira, Ana C. & Voss, Robert S. 2017 |
Monodelphis saci
| Pavan & Mendes-Oliveira & Voss 2017 |
M. pinocchio
| Pavan 2015 |
M. handleyi
| Solari 2007 |
M. handleyi
| Solari 2007 |
M. ronaldi
| Solari 2004 |
M. ronaldi
| Solari 2004 |
M. reigi
| Lew and Perez-Hernandez 2004 |
M. kunsi
| Pine 1975 |
M. kunsi
| Pine 1975 |
M. kunsi
| Pine 1975 |
M. kunsi
| Pine 1975 |
M. kunsi
| Pine 1975 |
M. kunsi
| Pine 1975 |
M. kunsi
| Pine 1975 |
M. kunsi
| Pine 1975 |