
Erythraeus ( Zaracarus ) budapestensis Fain & Ripka, 1998
|
publication ID |
https://doi.org/10.11646/zootaxa.5150.3.3 |
|
publication LSID |
lsid:zoobank.org:pub:EC7D1A70-3528-42A1-80D7-B57380CA5DC4 |
|
DOI |
https://doi.org/10.5281/zenodo.6645799 |
|
persistent identifier |
https://treatment.plazi.org/id/038C87FD-3D76-D703-D584-FF6EFB474EF0 |
|
treatment provided by |
Plazi |
|
scientific name |
Erythraeus ( Zaracarus ) budapestensis Fain & Ripka, 1998 |
| status |
|
Erythraeus ( Zaracarus) budapestensis Fain & Ripka, 1998 View in CoL
E. ( Z.) preciosus Goldarazena & Zhang, 1998 (synonymized by Haitlinger & Šundić 2015a).
E. ( Z.) iranicus Saboori & Akrami, 2001 syn. nov.
E. ( Z.) ueckermanni Saboori, Nowzari & Bagheri-Zenouz, 2004 (synonymized by Haitlinger & Šundić 2015a).
E. ( Z.) hamedamicus Khanjani, Miromayedi, Nahad & Fayaz, 2010 (synonymized by Mahmoudi et al. 2011 with E. ( Z.) ueckermanni).
Material examined. 2 deutonymphs reared from larvae sampled from Uroleucon ( Uromelon) jaceae (L.) and U. ( Uroleucon) chondrillae (Nevsky, 1929) (Aphididae) , Turkey, Erzincan province, 25 July to 15 August 2019; 3 adults collected manually from soil samples in Turkey, Erzincan province, 1 July 2020 , plus 14 larvae obtained from one female of collected adults.
Depository. Larvae collected as parasites have been lost. Vouchers representing all active instars and a remain of larvae freshly obtained from a female are deposited in the acarological collection at the Acarology Laboratory of Erzincan Binali Yıldırım University, Erzincan, Turkey ( EBYU) .
Diagnosis. Adult. Habitus typical for the genus, palp genu with 2–4 conalae and 2–3 semiconalae; palp tibia with 4–6 conalae and without semiconalae. Anterior sensillary area of crista metopica with 9–10 nonsensillary setae (AM). Anterior and posterior processes of crista metopica not detectable. Dorsal opisthosomal setae uniform of various lengths (20–90), covered sparsely with delicate setules. Distinct serratalae present on telofemur-tibia IV, on telofemora II–III weak, absent on genua and tibiae I–III.
Deutonymph. Similar to adults but much smaller ( Table 1 View TABLE 1 ). Palp genu with 0–2 conalae and 1–2 semiconalae, palp tibia with 3 conalae and without semiconalae. Anterior sensillary area of crista metopica with 5–8 nonsensillary setae (AM). Dorsal setae (16–75) similar to those of the adults. Genital sclerite small (26–30), without median opening.
Larva. Habitus as typical for the subgenus, AL inflated basally, ASens moderately short (30–35), bases of ASens setae with pouch-like structures. fD = 33–36, fV = 8–10, fnbFe = 2-2-2, length of tarsus III <120, length of tibia III 155–236.
Description. Adult ( Figs. 1–2a View FIGURE 1 View FIGURE 2 ) (based on 3 specimens). Metric data in Table 1 View TABLE 1 . Body length 1700–1935, width 1140–1200. Body reddish-dark brown and eyes bright red in live specimens.
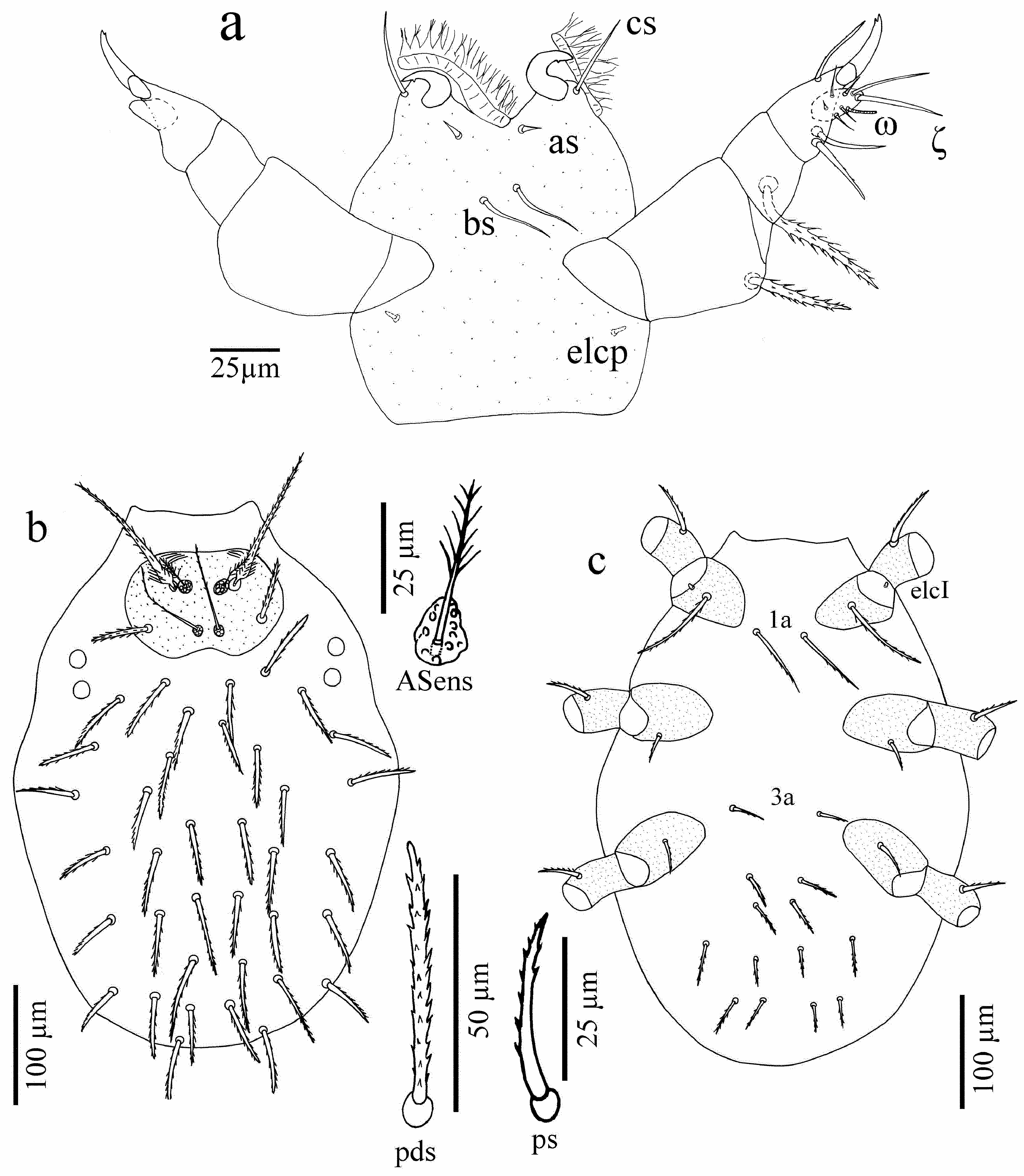
Gnathosoma. Chelicerae typical, of erythraeid type. Palps with relatively sparse setation. Normal, setulated setae present on palp trochanter and palp femur. Palp genu with strong, serrate setae dorsally, smooth and more slender setae laterally, with 2–4 conalae and 2–3 semiconalae. Palp tibia with normal, sparsely barbed setae, with 4–6 conalae. Odontus hook-like and with a small basal tooth ( Fig. 1a View FIGURE 1 ). Palp tarsus hemispherical, with the apical part overreaching the termination of odontus, densely covered with eupathidia and solenidia.
Idiosoma. Scutum ( Fig. 1c View FIGURE 1 ) clearly detectable and extending crista metopica laterally. Anterior and posterior processes of crista metopica not detectable in any specimen observed. ASA with 9–10 setulose, relatively long nonsensillary setae (AM) ( Fig. 1b View FIGURE 1 ). ASens with very short setules in its distal third. Bases of ASens with pouch-like structures. PSens with very short setules in their distal thirds. PSens slightly longer than Asens, bases of PSens with bowl-like cavity structures. Punctated ocular plates (60–75 × 80–88), located laterally of crista, anterior and posterior lens of about the same size (diameter: 26–28) ( Fig. 1c View FIGURE 1 ). Dorsal setae setiform, of various lengths (20–115). All dorsal setae originate from small plates. Stems are slightly narrowed terminally and covered sparsely with delicate setules ( Fig. 1d View FIGURE 1 ). Ventral setae (35–65) thinner, covered with setules throughout its 4/5 of length ( Fig. 1e View FIGURE 1 ). GOP surrounded by paired sclerites (genital plates: epivalves with up to 60 setae which are smooth or with very short barbs and centrovalves without setae), located at the level of coxae III–IV ( Fig. 1f View FIGURE 1 ). AOP markedly shorter than genital opening, surrounded by paired anal valves of which the external ones are stronger sclerotized and bear 4–5 pairs thin and short (20–35) setae with very short barbs ( Fig. 1g View FIGURE 1 ).
Legs. Legs IV much longer than legs I–III (legs II shortest). Serratalae ( Fig. 2a View FIGURE 2 ) on telofemora II–III weak, absent on genua and tibiae I–III. Normal setae with small setulae present on all leg segments. Tarsi I–IV terminated in paired claws, each claw covered with fimbriae.
Deutonymph ( Figs. 2b–e View FIGURE 2 ) (based on 2 specimens). Metric data in Table 1 View TABLE 1 . Similar to adults but much smaller, body length 620–890, width 380–720. Body reddish-dark brown in live specimens. Palp genu with 0–2 conalae and 1–2 semiconalae, palp tibia with 3 conalae ( Fig. 2b View FIGURE 2 ). Scutum clearly detectable and laterally extending to level of ASA of crista metopica. Anterior sensillary area of crista metopica with 5–8 nonsensillary setae (AM) ( Fig. 2c View FIGURE 2 ). Dorsal setae (16–75) similar to those of the adults. Genital sclerite small (26–30) carrying a pair of smooth, thin setae; and without opening ( Fig. 2d View FIGURE 2 ). Anal sclerites surrounding the anal opening elongate (55–59) and with 3 pairs of smooth, thin setae ( Fig. 2e View FIGURE 2 ).
Larva ( Figs. 3–6 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 ) (based on 14 specimens). Metric and meristic data in Tables 2–3 View TABLE 2 View TABLE 3 . Body length 460–755, width 275–510 ( Fig. 3 View FIGURE 3 ). Body orange in live specimens.
Gnathosoma. Cheliceral blades with a small subterminal tooth. Dorsal gnathosoma with a pair of smooth, pointed adoral setae (cs) (30–32) anteriorly and a pair of minute supracoxal setae (elcp) (5–7) in lateral position. Ventrally a pair of smooth, pointed subcapitular (tritorostral) setae (bs) (30–38) and a pair of spine-like oral setae (as) (9–10). Palp femur and genu bear one setulose seta dorsally, seta (35–45) on palp femur always shorter than seta (48–60) on palp genu. Palp tibia with three setae: one nude and two barbed setae. Palp tibial claws bifid in its distal half. Palp tarsus with one solenidion, one eupathidium, five nude setae and a tiny spine-like seta ( Fig. 4a View FIGURE 4 ). Palp formula: 0-N-N-NNN 2 -NNNNNNωζ.
Idiosoma ( Figs 4b–c View FIGURE 4 ). Scutum punctate, oval and wider than long, anterior border straight, posterior border slightly concave, lateral borders convex. ASens and PSens with barbs in its distal half, PSens much longer than ASens. Bases of both sensillary setae with pouch-like structures (detail in Fig. 4b View FIGURE 4 ). Anterior normal setae (AL) setulose and widened near basis. Posterior normal setae (PL) setulose, half the length of AL. Two pairs of eyes located postero-laterally to scutum, lenses are almost equal in diameter (ca. 14). Dorsal setae of various lengths (42–70), setulose. The total number of dorsal and ventral setae (NDV) ranges from 42 to 46, not considering setae located on scutum and coxae as well as intercoxal (1a, 3a) setae ( Figs. 4b–c View FIGURE 4 ). Coxa I with short (4–5) supracoxal seta (elcI) and long (75–98), serrate seta 1b. Serrate seta 1a (65–77) placed outside just behind the posterior border of the coxal plate I. Coxa II with short (28–36), serrate seta 2b. Serrate seta 3a (30–40) placed outside at anterior border of coxal plate III. Coxa III with serrate seta 3b (32–44). Posterior to coxae III four pseudanal setae (25–45), which are slightly thinner than the neighbouring idiosomal setae located more posteriorly ( Fig. 4c View FIGURE 4 ).
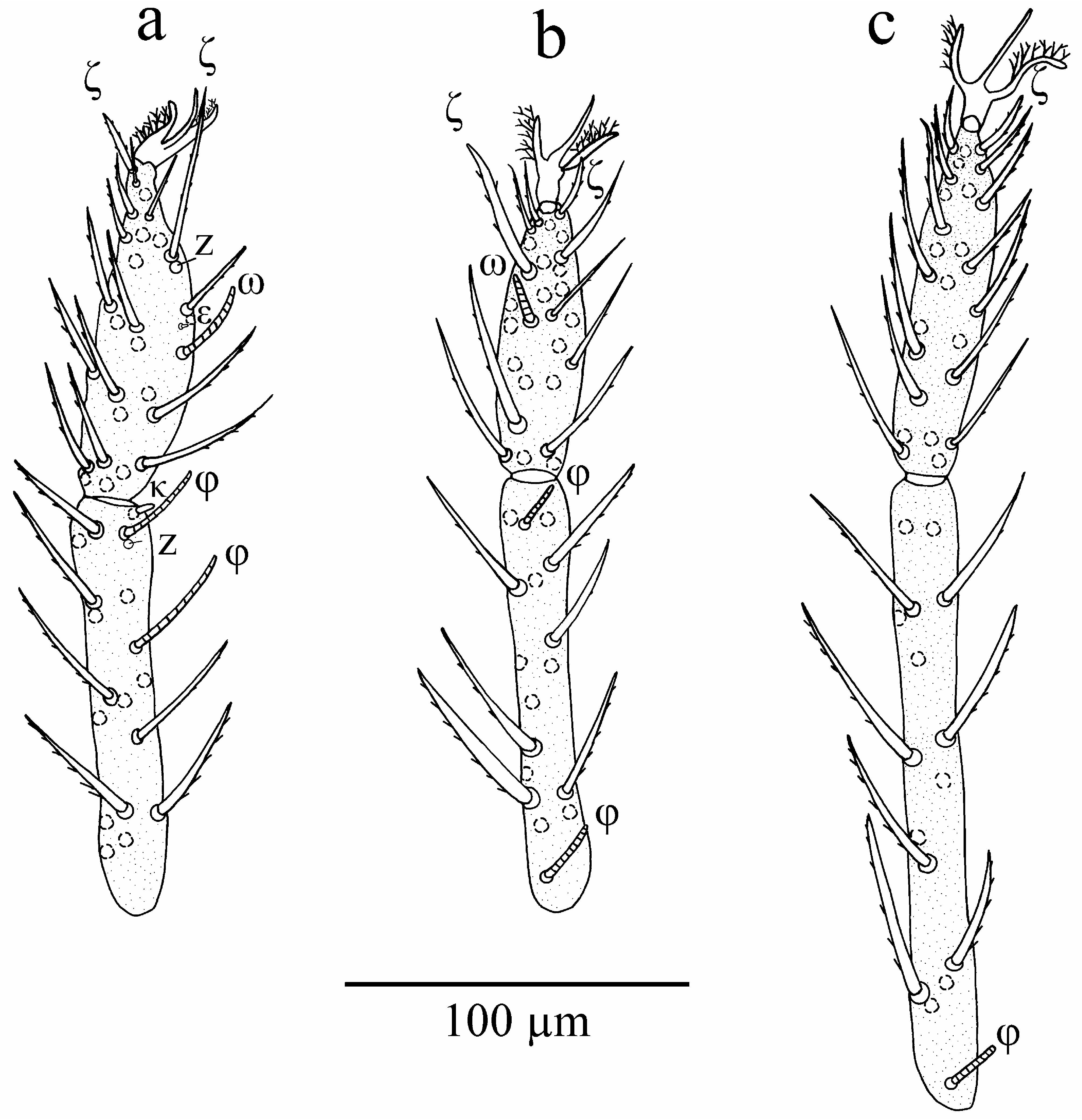
Legs ( Figs. 5–6 View FIGURE 5 View FIGURE 6 ). Segmentation formula: 7-7-7. For leg chaetotaxy and morphometric data see Tables 2–3 View TABLE 2 View TABLE 3 . Solenidia on genu I and tibia I small (20–40), both solenidia (φ) on tibia I in distal half of the segment, distal solenidion on tibia I with accompanying seta z. Solenidion (ω) on tarsus I prominent (25–35), famulus (ε) small (3–4), inconspicuous. Subterminal eupathidium on tarsus I long (40–50) and with accompanying seta z (6–8), ‘pretarsal’ eupathidium shorter (15–20), without seta z. All solenidia on leg II short (14–20); solenidia on tibia II close to the distal and proximal border of the segment, respectively. Subterminal eupathidium on tarsus II long (36–44), no accompanying seta z detected, ‘pretarsal’ eupathidium shorter (14–20). Solenidion on tibia III short (15–20) and close to the proximal border of the segment, ‘pretarsal’ eupathidium short (16–20). Median empodium of pretarsi I–III claw-like, with small lateral barbs. Anterior lateral claw with, posterior lateral claw without terminal hook. Both lateral claws with several branching setules (aliform).
Distribution. Albania, Bosnia and Hercegovina, Bulgaria, Croatia, Greece, Hungary, Iran, Italy, Montenegro, Serbia, Spain, Turkey and Ukraine ( Saboori & Akrami 2001; Mąkol & Wohltmann 2012, 2013; Haitlinger & Šundić 2015a, Haitlinger & Mehrnejad 2017).
Taxonomic remarks. Of the 28 species described in the subgenus Zaracarus ( Xu et al. 2019) , 17 species belong to the group of species having three normal setae on basifemora I–II, while the remaining 11 species are characterized by having two normal setae on basifemora I–III. Of the latter group, six species are characterized by the base of seta AL being swollen, setulose: E. ( Z.) budapestensis ; E. ( Z.) iranicus ; E. ( Z.) hamedamicus; E. ( Z.) eleonorae Haitlinger, 1987 ; E. ( Z.) arminouensis Haitlinger & Łupicki, 2011; E. ( Z.) kurdistaniensis Khanjani & Ueckermann, 2005 and E. ( Z.) didonae Haitlinger 2000 . In the key published by Xu et al. (2019), the latter species was erroneously included into the group of species having 3 normal setae on basifemora I–III.
We did not detect convincing differences between larvae of E. ( Z.) budapestensis and E. ( Z.) iranicus , which was originally described from a single specimen from Iran ( Saboori & Akrami 2001) and further supplemented by the additional analysis of larvae collected in Turkey ( Oner et al. 2021). Based on the larger number of specimens, we found overlapping of metric data in these species. Moreover, the number of normal setae on tibiae I–III is not as stable as assumed by Saboori & Akrami (2001) and Noei et al. (2017). Haitlinger & Šundić (2015a) reported 14–15 setae on each tibia of legs I–III based on checking 47 larvae determined as E. ( Z.) budapestensis and collected in different countries. The differences of the number of normal setae on tibiae II–III found in the present investigation confirm that such variability is also present within a particular population. Differences in the number of normal setae do not certainly indicate species borders. This is also supported by the variability in the number of normal setae on tibiae found in Erythraeus larvae emerged from eggs deposited by a single female in the laboratory ( Stålstedt et al. 2016). The only other difference between E. ( Z.) budapestensis and E. ( Z.) iranicus refers to the number of normal setae on palp tarsus, a character which is often difficult to detect in Erythraeus larvae. For these reasons, we consider E. ( Z.) iranicus as subjective junior synonym of E. ( Z.) budapestensis .
The statement of Saboori & Shirvani (2021) that the number of normal setae on tibiae is stable and does not vary in re-investigated specimens of E. ( Z.) hamedamicus does not justify re-establishing of its species status considering the variability found in E. ( Z.) budapestensis . Thus, the synonymization of E. ( Z.) hamedamicus with E. ( Z.) ueckermanni by Mahmoudi et al. (2011) was correct. Since E. ( Z.) ueckermanni was synonymized with E. ( Z.) budapestensis by Haitlinger & Šundić (2015a), E. ( Z.) hamedamicus must also be regarded as subjective junior synonym of E. ( Z.) budapestensis .
From the remaining species of the group with two normal setae on basifemora I–III, E. ( Z.) budapestensis differs in having the shorter genu II (<120 vs>120) and smaller width of scutum (max 150 vs min 155) (see also taxonomic discussion for E. ( Z.) rupestris ).
The deutonymphs and adults of E. ( Z.) budapestensis fit the general set of characters of Erythraeus species known from postlarval instars, and do neither match the particular characteristics of the subgenus Parerythraeus nor display obvious differences from postlarval instars assigned to the nominate subgenus Erythraeus . Further differentiation is limited by the fact that many descriptions of postlarval instars have been published relatively long ago and do not match present taxonomic standards, and only a few mites, mostly belonging to European species, have recently been redescribed ( Gabryś 2016a, b; Stålstedt et al. 2016). No postlarval stages of Erythraeus species have hitherto been reported from Turkey, and comparison of E. ( Z.) budapestensis deutonymphs and adults to published species descriptions resulted in no match.
Of course we cannot exclude that more than only one species is hidden in the pool of specimens summarized as E. ( Z.) budapestensis . The current way of species descriptions based on single or few specimens representing only a particular instar has obvious limitations. Further research to combine different ontogenetic instars, to include life history features and molecular characters will certainly help to resolve the problems of species diagnostics.
Biology. Field data. Larvae ectoparasitic on aphids, Uroleucon ( Uromelon) jaceae (Linnaeus, 1758) and U. ( Uroleucon) chondrillae (Nevsky, 1929) , were sampled using sweep nets in the period from July 25 to August 15 in 2019. These samples were collected from three different locations in Erzincan province of Turkey ( 39°47’ N, 39°28’ E, Işıkpınar village, 1332 m a.s.l.; 39° 47’ N, 39° 27’ E, Demirkent village, 1257 m a.s.l; and 39° 38’ N, 39° 31’ E, Yaylabaşı village, 1278 m a.s.l.). All locations are semi-aquatic grassland. Of about 1100 Uroleucon specimens observed at these locations, 60 Uroleucon specimens (ca. 5%) were parasitized by E. ( Z.) budapestensis larvae. Of them, 41 hosts were parasitized by one E. ( Z.) budapestensis larva, 16 were parasitized by two, two—by three, and one—by four mite larvae. A total of 83 mite larvae were counted on hosts; among them, 73 larvae were attached to the head (between the antennae), the remaining were attached to the thorax (4), abdomen (3) and legs (3) of Uroleucon aphids ( Fig. 7 View FIGURE 7 ).
Three adults were collected manually from the soil surface on 1 July 2020 in Erzincan Province, Turkey, 39° 44’ N, 39° 30’ E, University campus, 1220 m a.s.l., in a semi-aquatic grassland area GoogleMaps .
Laboratory data. Of nine larvae collected from Uroleucon aphids, six deutonymphs emerged from protonymphs 6–12 days after placing the fed larvae in glass vials. Freshly emerged deutonymphs were preserved for taxonomic study. Of the three adults collected in early July 2020, one female deposited about 50–60 black eggs in a cluster three days after capturing. In total, 42 larvae hatched within 23 days after oviposition.
Discussion on Biology. Since the correspondence of larval and postlarval instars was the general aim of laboratory rearing, duration of instars was not a special focus of the study. However, development data fit the sequence of instars known from other Erythraeus species ( Stålstedt et al. 2016) and Parasitengona in general ( Wohltmann 2000). With regard to the life cycle of E. ( Z.) budapestensis , it remains unknown whether a diapause is present and which instar might be the hibernating one. However, neither egg nor protonymph interrupt the continuous development in this species. Published host reports for E. ( Z.) budapestensis refer exclusively to Aphidae but are not restricted to the genus Uroleucon ( Saboori & Akrami 2001; Haitlinger & Mehrnejad 2017; Oner et al. 2021).





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Erythraeinae |
|
Genus |
|
|
SubGenus |
Zaracarus |