
Bemisia flocculosa, Gill, Raymond & Holder, Peter, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.206893 |
|
DOI |
https://doi.org/10.5281/zenodo.6185221 |
|
persistent identifier |
https://treatment.plazi.org/id/038D87CF-FFFB-4320-059F-69A9FDD2FEEC |
|
treatment provided by |
Plazi |
|
scientific name |
Bemisia flocculosa |
| status |
sp. nov. |
Bemisia flocculosa sp. n.
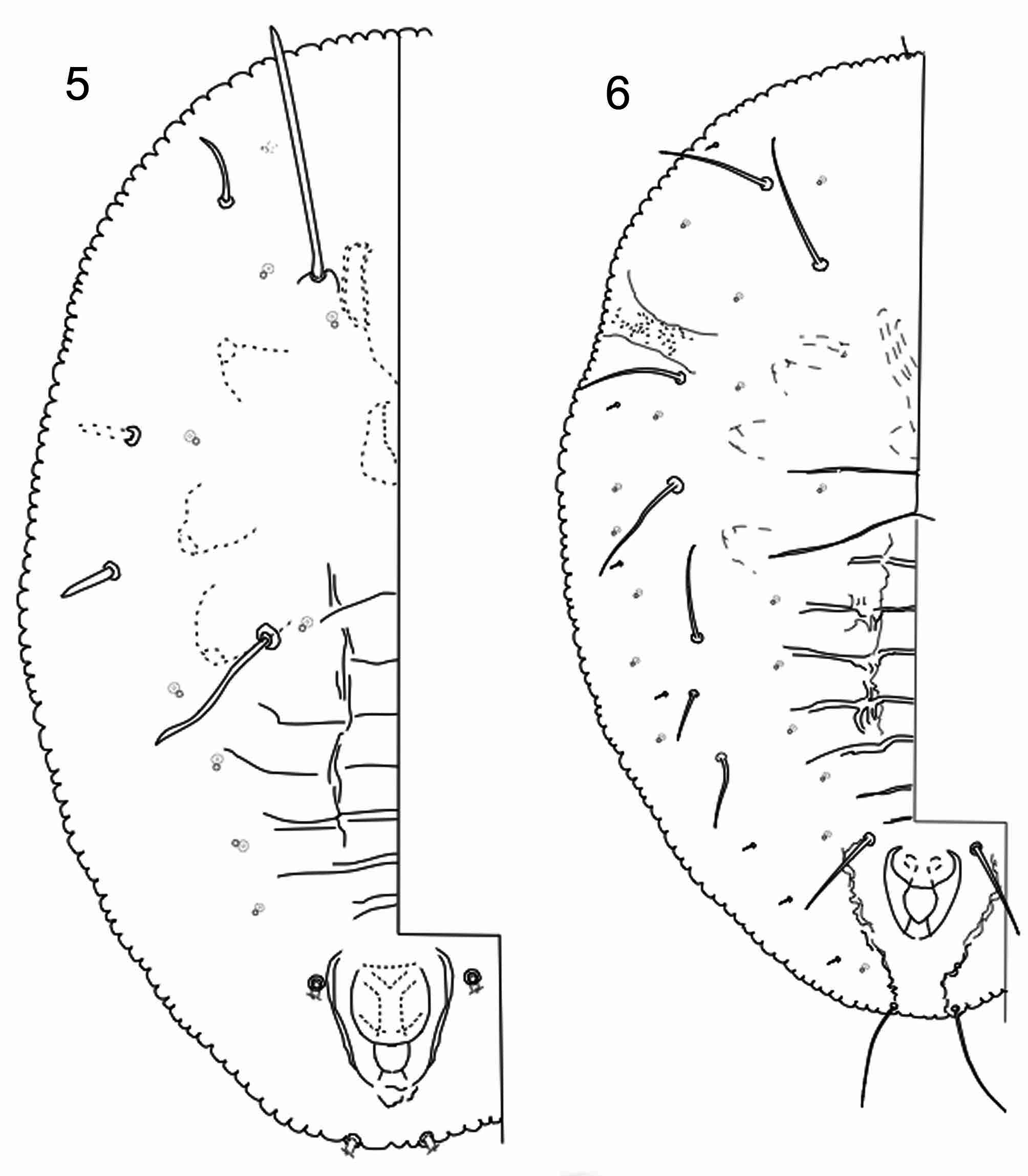
Puparium (4th instar nymph; Figs. 1 View FIGURES 1 – 4 & 6 View FIGURES 5 – 6 )
Habitus: Oval or elliptical, widest at a point opposite meso/meta-thoracic suture. Dimensions variable; length and width of holotype 830 microns long by 620 microns wide; paratype specimens 810–1020 microns long (avg. 930 microns) by 580–790 microns wide (avg. 704 microns). Body flat, color light yellowish brown. Dorsum with noticeable wax secretions consisting of white flocculent wax in a row on each side between the dorsal disk and the inner edge of the submargin and extending from the head posteriorly as far as the vasiform orifice, and a single row along the abdominal midline and thorax. The dorsal wax secretions appear serrated due to the placement along the segmental divisions, less wax appearing at the division lines. In addition to the flocculent wax, seta-like transparent wax rods are produced around the outside margin, presumably formed from the submarginal pore/porette combinations in those locations.
Morphology. Margin with shallow irregular crenulations, numbering about 125 per side on the type specimen. Differentiated crenulations at the thoracic tracheal furrows absent; crenulations in the caudal area barely distinct from the others but generally a bit smaller. Ventral surface thoracic tracheal furrows with scattered, numerous microspines. Marginal seta in the usual locations anteriorly and posteriorly but very tiny (7–12 microns long) and often broken off during the slide mounting process. Dorsal disc flat except for raised callosities and wax producing areas occurring in a narrow arc from the cephalic area to roughly the 7th abdominal segment, and with centrally located raised wax-producing areas on the 2nd to 7th abdominal segments along the midline and paired elevations centrally on the thoracic segments. Submargin poorly delineated, but reaching the raised callosities and wax producing arcs delineating the outer edges of the dorsal disk. Transverse molting suture reaching but not surpassing discal callosity arc; longitudinal molting suture terminating before anterior margin, with short branches laterally at the apex. Dorsal pore/porette combinations are adjacent, with no particular alignment with the margins of the pupal case. Pores on the dorsal disc forming two single pore lines on each side of abdominal segments and also forming a sparse single band around the submargin, otherwise occurring randomly. First abdominal segment with one pair of pore/porettes on dorsal disc area. Dorsal setae consist of pairs of usually short setae on the disc and on the submargin. Most dorsal setae short, 7–12 microns long, except for the 8th abdominal setae (60–90 microns long, avg. 73.5 microns) and the caudal setae (48–78 microns long, avg. 60 microns). Occasionally, a dorsal seta may elongate to 50–66 microns long, either on the cephalothorax or the 4th abdominal segment, although usually only one seta will enlarge on any given specimen. Five pairs of dorsal disc setae along outer edge of callosity arc at edge of submarginal area, three pairs on cephalothorax and two pairs on abdomen including 8th abdominal pair; two pairs centrally on disk, one pair in area of prothorax and one pair in 1st abdominal segment; with one pair of caudal setae near posterior margin. With 9–10 pairs of shorter submarginal setae located in an arc midway between margin and callosity arc and dorsal disc. Vasiform orifice ovate to chordate, open posteriorly; operculum broadly rounded posteriorly, not totally covering lingula and covering about one half of orifice. Posterior sides of orifice with strengthening ridges but no indications of teeth or similar structures. Lingula spatulate, tapering gradually posteriorly, with a pair of setae distally.
Third instar ( Fig. 6 View FIGURES 5 – 6 )
Habitus: Length of body (three individuals) 570–630 microns (avg. 608 microns); width 420–462 microns (avg. 438 microns). Field appearance unknown.
Morphology: Margin with shallow crenulations, numbering about 96–100 per side. Differentiated crenulations not apparent at ends of tracheal furrows, but those in the caudal area appear shallower. Ventral area of thoracic tracheal furrows with scattering of microspines. Marginal seta in the usual locations anteriorly and posteriorly. With 7–8 pairs of enlarged dorsal setae, lengths 60–90 microns on each individual. Submargin not delineated from dorsal disc except that the enlarged dorsal setae appear to align with the discal/submarginal arc. With 7–8 pairs of short, minute submarginal setae aligned in a single row midway between the body margin and edge of the apparent dorsal disc. Submarginal pores of simple pore/porette type sparse, in apparently two incomplete rows. Vasiform orifice chordate, lingula spatulate, similar to that of the adult.
Second instar ( Fig. 5 View FIGURES 5 – 6 )
Habitus: Unknown.
Morphology: Measurements 468 microns long by 300 microns wide (based on one specimen only, a 2nd/3rd instar molt, with the 3rd instar nearly fully formed inside of 2nd). The single specimen with four pairs of enlarged dorsal setae on cephalothorax, three setae (one pair completely broken off at setal base) on the submarginal/ discal area, and one longer setal pair in a more median location of the prothoracic area above the mouthparts. A dorsal discal pair of elongate setae on first abdominal segment, a setal pair each on the eighth abdominal segment lateral to the vasiform orifice and pair of caudal setae (both pairs broken off completely at setal base). Marginal and submarginal setal positions not discernable. Discal pore/porette groups in a single row on four abdominal segments and randomly scattered on cephalothorax. Locations of submarginal pore/porettes not clear due to presence of same pore groups on 3rd instar underneath. Vasiform orifice chordate, operculum not filling orifice or totally covering lingula. Lingula roundly and broadly spatulate.
First instar. Unknown.
Adult ( Figs 2–4 View FIGURES 1 – 4 )
Morphology: Specimens typical of Aleurodine adults, with no outstanding or unique morphological structures. Dimensions of slide-mounted specimens: males 1.10–1.30mm long (avg. 1.18mm), females 1.20–1.42mm (avg. 1.27 mm). Dimensions of hind tibiae: males 348–414 microns long (avg. 385 microns), females 420–462 microns (avg. 433 microns); length of antennae: males 228–330 microns (avg. 286 microns), females 280–318 microns (avg. 305 microns); length of third antennal segment: males 120–132 microns (avg. 126 microns), females 126– 142 microns (avg. 139 microns). Upper and lower compound eyes connected by multiple ommatidia, two on one side against two or three on the other. Length of male claspers (parameres) 138–150 microns (avg. 144 microns). Clasper without subapical teeth or other distinguishing morphology. Metatibial comb containing 18–20 setae, not quite reaching metatibial brush apically. Dorsum of male last abdominal segment with 3 pairs of small setae in lines lateral to and posterior to anal opening and lingula.
Material examined. Holotype puparium second specimen from left on slide with 5 other puparia, marked with black lines: NEW ZEALAND, MC., Christchurch, Botanical Gardens Natives Section, Site 2, NPPRL Ref. 3/ 2004/1050, on Melicytus aff. obovatus leaf, 19.iii.2004 (P. Holder/M. Bullians), deposited in New Zealand Arthropod Collection, Landcare Research, Auckland, New Zealand.
Paratypes, all NEW ZEALAND, MC, Christchurch Botanical Gardens: one slide, same data as holotype except, Ref. 3/2004/1049, with 5 pupae & 1 3rd instar nymph; one slide, same data as holotype except, Ref. 3/2004/ 1048, with 9 pupae; one slide (partially handwritten label), 5 pupae, Site 10, NPPRL Ref. 3/2004/1048, on Melicytus obovatus , 19.iii.2004, no collector listed; one slide (partially handwritten label), 8 pupae, Site 5, NPPRL Ref. 3/ 2004/1046, on Melicytus lanceolata , 19.iii.2004 (P. Holder); one slide, 6 pupae, Site 9, NPPRL Ref. 3/2004/1048, on Melicytus obovatus , 19.iii.2004 (P. Holder/M. Bullians); one slide, 6 pupae, Site 1, NPPRL Ref. 3/2004/1085, on Melicytus alpinus , 19.iii.2004 (P. Holder/M. Bullians); one slide, 6 pupae, Site 11, NPPRL Ref. 3/2004/1048, on Melicytus obovatus , 19.iii.2004 (P. Holder/M. Bullians); one slide, 6 pupae, Site 7, NPPRL Ref. 3/2004/1048, on Melicytus obovatus , 19.iii.2004 (P. Holder/M. Bullians); one slide, 6 pupae, Site 5, NPPRL Ref. 3/2004/1046, on Melicytus lanceolatus , 19.iii.2004 (P. Holder/M. Bullians); one slide, 2 adult males, 1 adult female, ex. Melicytus obovatus , 30.vi2008 (Peter Holder); one slide, 2 adult males, 2 adult females, 3 pupae, ex. Melicytus obovatus , 30.vi.2008 (Peter Holder); one slide, 2 adult males, 2 adult females, 2 pupae, ex. Melicytus obovatus , 30.vi.2008 (Peter Holder); one slide, 1 adult male, 1 adult female, ex. Melicytus obovatus , 30.vi.2008 (Peter Holder); numerous unmounted dry specimens on original leaves, ex. Melicytus obovatus , 30.vi.2008 (Peter Holder).
Paratypes will be distributed to: California Department of Food and Agriculture, Sacramento California, U.S. National Museum, Beltsville, Maryland, The Natural History Museum, London, and New Zealand Arthropod Collection, Landcare Research, Auckland, New Zealand.
Biology. In the Christchurch population observed, B. flocculosa appears to be primarily univoltine, or partially bivoltine. Adults have only been seen for one six week period in late spring, from early November to mid-December on Melicytus obovatus , and approximately a week longer on M. ramiflorus . The population is highly seasonally synchronized, with nymphal life stages developing slowly, as a single cohort, over summer, winter and spring. Two distinct puparial sizes were observed. This may be a gender difference, or represent dispersive versus non-dispersive individuals. Close observation revealed that newly molted third instar nymphs were capable of rotating about their stylets.
This new species has been collected from six Melicytus species: M. obovatus ; M. alpinus ; M. micranthus ; M. lanceolatus ; M. flexuosus ; M. ramiflorus ; M. aff. obovatus (= sp. 1 and M. obovatus ).
Distribution. This new species is so far confirmed only from Mid-Canterbury, New Zealand. In addition to the Christchurch Botanic Gardens collection records presented here, other non-paratype collections have been found on preserved and modern Melicytus micranthus at Riccarton Bush (coll. in 1957 [A.J. Healy] and 2007) and Prices Valley, Banks Peninsular (coll. 2005 (later collections N.A. Martin)). This additional collection material is housed at the New Zealand Arthropod Collection, Landcare Research, Auckland, New Zealand.
Bemisia flocculosa has been morphologically matched with samples in the Natural History Museum, London, collected in Israel in 1986, on Melicytus sp., probably M. ramiflorus . As the host plant is a New Zealand endemic, it is concluded that the whitefly has most likely been dispersed to Israel from New Zealand with the host material (John Martin pers comm. April 2004).

| NEW |
University of Newcastle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |