
Pogonostoma (Pogonostoma) meridionale Fleutiaux, 1899
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4790.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:6316903F-9894-40CF-A22C-8593637F1C9A |
|
persistent identifier |
https://treatment.plazi.org/id/038E87CB-D313-CD07-FF21-FDC58736B27D |
|
treatment provided by |
Plazi |
|
scientific name |
Pogonostoma (Pogonostoma) meridionale Fleutiaux, 1899 |
| status |
|
Pogonostoma (Pogonostoma) meridionale Fleutiaux, 1899 View in CoL
( Figs 27–51 View FIGURES 19–32 View FIGURES 33–41 View FIGURES 42–47 View FIGURES 48–51 ; 52 View FIGURE 52 )
Pogonostoma meridionale Fleutiaux, 1899: 384 View in CoL .
Type locality. Fort Dauphin (now Tolagnaro), south-eastern Madagascar .
Pogonostoma (Pogonostoma) meridionale: Jeannel 1946: 123 View in CoL .
Pogonostoma (Pogonostoma) meridionale: Rivalier 1970: 304 View in CoL .
Pogonostoma (Pogonostoma) meridionale: Moravec 2007: 291 View in CoL .
Type material. Lectotype ♂ (designated by Moravec 2007) in MNHN (coll. Générale), labelled: “Fort Dau- phin” [handwritten] // “Museum Paris / Coll. E. Fleutiaux” [blue, printed] // “ Pogonostoma meridionale Fleut. type / Coll. Fleutiaux” [handwritten/printed] // “Type” [red, printed] // “ Lectotype / Pogonostoma / meridionale Fleutiaux, 1899 / design. Jiří Moravec 2003” [red, printed]. Paralectotype. 1 ♀ in MNHN (Collection Jeannel) with same first two labels.
Other material examined. Historical data. 1 ♀ in MNHN (Collection Jeannel) with same labels as paralecto- type. 8 spms (♂♂, ♀♀) in MNHN (Coll. Jeannel) : “ Bas Mandrare Ampasimpolaka ”. 6 spms (♂♂, ♀♀) in MNHN (Coll. Jeannel) : “ Basin du Mandrare ” . 1 ♂, 1 ♀ in MNHN , 2 ♂♂, 2 ♀♀ in BMNH : “ Plateau de l’ Androy, reg. d’Ambovombe” . 2 ♂♂, 2 ♀♀ in SDEI : “ Andrahomana ” . 1 ♂, 1 ♀ in BMNH , 24 spms (♂♂, ♀♀) in MNHN , 1 ♂, 1 ♀ in CCJM (ex APCA) : “ Behara ”. 20 spms in MNHN (partly in Coll. Jeannel) , 24 spms (♂♂, ♀♀) in SDEI , 4 ♂♂, 1 ♀ in MFNB , 1 ♂ in CCJM (ex APCA) : “ Bekily ” . 1 ♂ in MNHN , 2 ♂♂, 1 ♀ in SDEI , 1 ♀ in MFNB , 1 ♂ in BMNH , 1 ♂ in CCJM (ex APCA) : “ Fort Dauphin ” . 1 ♀ in SDEI , 4 ♂♂ in MFNB , 3 ♂♂ in NHMW : “ Madagascar ”. 1 ♂ in SDEI : “ Annanarivo ” .
Recent data. 1 ♂ in CCJM “ Madagascar / Fort Dauphin / 24 km W / 11.II.2006 / A. Monfort Col. ” .
Differential diagnosis. Pogonostoma (P.) meridionale is immediately distinguished from other species of the Pogonostoma (P.) elegans species-group by its ferrugineous setal vesture which is particularly conspicuous on elytra while on the head and pronotum the setae are better obvious in lateral view (only in very old, badly treated specimens the rusty setae fade). The setae in the similar P. (P.) natsuae sp. nov. are generally paler, blonde to straw-yellow. The aedeagus of P. (P.) meridionale (in its left lateral aspect, Figs 28, 30 View FIGURES 19–32 ) is notably boomerang-bent. Moreover, the aedeagus clearly differs from that in P. (P.) natsuae sp. nov. in the shape of its apex, particularly in its ventral (and dorsal) aspect ( Figs 29, 31, 32 View FIGURES 19–32 ). For other differences see “Differential diagnosis” under the new species above.
P. (P.) rivalieri Moravec, 2005 may resemble P. (P.) meridionale by its black body, subgibbose pronotal disc in its lateral aspect, nearly smooth pronotal posterior lobe, partly reddish-brown mandibles and similar aedeagi. However, P. (P.) rivalieri is immediately distinguished (both from P. (P.) meridionale and P. (P.) natsuae sp. nov.) by its elytral sculpture consisting of much larger, deeper and more regular punctures and by the shape of its pronotal disc (particularly in male), which is subquadrate in dorsal view (see Moravec 2005, 2007). Another rather similar species, P. (P.) atrorotundatum W. Horn, 1934, clearly differs in having elytral sculpture consisting of extremely large punctures with thin intervals and its pronotal posterior lobe being punctate-setose.
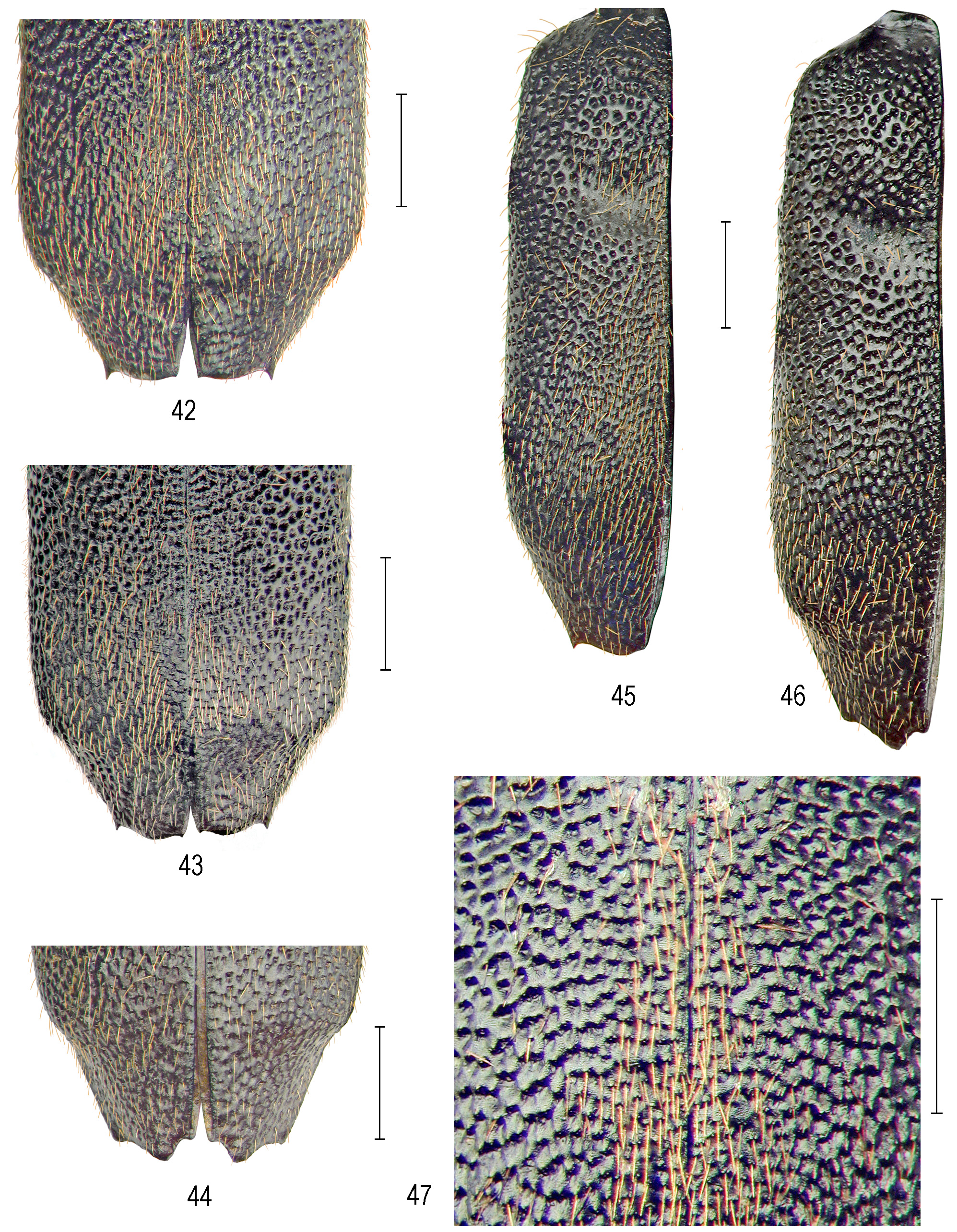
Apart from the distinct ferrugineous setal vesture, P. (P.) meridionale clearly differs from the above-mentioned species in having elytral sculpture of various patterns ( Figs 42–47 View FIGURES 42–47 ) formed by much denser punctures with flat, matt and multiform intervals, densest and irregular along sutures and with distinctly aciculate anterior margins on posterior elytral surface ( Figs 42–44 View FIGURES 42–47 ).
For other differences from the above-mentioned species and from others of the P. (P.) elegans species-group see Moravec (2007).
Redescription. Body ( Fig. 33 View FIGURES 33–41 ) 10.1–12.5 (LT 11.0) mm long, 2.60–3.30 (LT 3.00) mm wide, black, matt; setal vesture ochre-ferrugineous.
Head ( Fig. 35 View FIGURES 33–41 ) slightly narrower than body, width 2.30–2.70 mm; temples moderately long (2 times shorter than eyes).
Frons and vertex shaped and with surface generally as in preceding species, but posterior impression very faint or absent, dorsal surface densely covered with erect, very short, ochre hairlike setae.
Genae and clypeus as in preceding species.
Labrum primarily 6-setose, rarely 5–8-setose with 3–5 (rarely 6) anterior and 2 lateral, dark-brown or reddishbrown setae, black, often with mahogany tinge particularly on basomedian area, surface generally as in preceding new species; male labrum ( Figs 38–39 View FIGURES 33–41 ) comparatively long, length 0.70–0.85 mm, width 1.30–1.40 mm, female labrum ( Figs 40–41 View FIGURES 33–41 ) more prolonged anteriad, length 0.90–1.20 mm, width 1.40–1.50 mm.
Maxillae ( Fig. 34 View FIGURES 33–41 ): galea black, usually with brownish apex of terminal galeomere, lacinia black-brown to reddish-brown (faded to rusty-brown in old specimens) with brownish or ochre-testaceous setae, apical portion with dilated outer margin, forming spatulate-dilated apex, 0.35–0.45 mm wide.
Palpi of usual length and shape, black except for cinnamon-brown inner apical area of terminal palpomeres, covered with scattered short whitish or greyish microtrichia, longest palpomeres of labial palpi with long, blackviolet setae.
Mandibles ( Figs 36–37 View FIGURES 33–41 ) with rather short terminal teeth, sexually dimorphic in shape as in preceding species, but generally much paler, dark reddish-brown to conspicuously reddish, or cinnamon-testaceous, except for sharply delimited, black-brown darkened basomedian area.
Antennae in male longer than body, in female slightly shorter; scape either black or cinnamon-brown, covered with sparse whitish microsetae and with subanterior easily abraded brownish seta, antennomeres 2–4 black, with scattered whitish microsetae, 5–11 with greyish-rusty microtrichia.
Thorax. Pronotum ( Figs 48–51 View FIGURES 48–51 ) elongate, length 2.55–3.20 mm, width 1.75–2.00 mm; shape and surface generally as in preceding new species, but notopleural sutures more notable near posterior discal angles in dorsal view and more distinctly subgibbose in lateral view (particularly in male), and the cristate-tuberculate sculpture consisting of generally larger tubercles and crests; whole discal surface covered with scattered, short, decumbent or erect, ochre or ochre-ferrugineous hairlike setae (better obvious in lateral view); lateral and ventral thoracic sterna generally as in preceding new species.
Elytra ( Figs. 42–47 View FIGURES 42–47 ) elongate, 5.70–7.50 mm long; humeri arcuate; lateral margins subparallel in male, moder- ately enlarged towards rounded lateral angles of anteapical convexity; elytral apices mostly notably sexually dimorphic: apex in male with mostly very faint sutural emargination towards indistinct sutural spine, arcuate in middle between small, thorn-like external tooth; rarely mesially forming indistinct, obtuse inner tooth; apex in female with usually deeper and steep sutural emargination and protruding, right-angled or rarely acute-angled inner tooth, and obtuse-angled, or more distinct, right-angled external tooth; elytral surface convex, basodiscal convexity moderate to distinct, discal impression deep and wide, surface notably multiform-punctate except for effaced narrow basal and basohumeral area; sculpture formed by punctures of various shape; they are deeper and larger and irregularly anastomosing with declined lateral intervals on basodiscal convexity, deep and more regular on lateral areas, rather large but commonly irregularly anastomosing on elytral disc, slightly reduced or entirely effaced within discal impression, very fine and irregular on wide discal area along sutures and expanded posteriad; smaller and spaced punctures with distinct aciculate anterior margins of their intervals cover the area of posterior declivity and whole anteapical and apical area, very irregularly anastomosing also on sublateral areas forming very irregular, multiform, flat intervals with aciculate anterior margins; surface of intervals matt, due to rough meshes of cuticular microsculpture; setal vesture consisting of nearly erect, copious, moderately long, notably ochre-ferrugineous or deep rusty ornamental setae which are mixed with sparse, long and erect, hairlike sensory setae.
Abdomen. Ventrites black with scattered, short, white setae.
Legs black (black-brown with mahogany-brown femora in old specimens); coxae rather densely whitish to yel- lowish-setose; profemora covered with rather dense medium-long whitish setae, mesofemora with dense whitish setae anteriorly, metafemora with sparser ochre and dark ferrugineous setae; pro- and mesotibiae with whitish or blonde setae except for dense, ochre-ferrugineous setae on apices of tibiae, tarsi dark reddish-brown their surface with whitish setae (as in all species, first three protarsomeres in male dilated and with pad of setae), claws rustybrown.
Aedeagus in its left lateral aspect ( Figs 28, 30 View FIGURES 19–32 ) conspicuously boomerang-shaped, 2.60-2.80 mm long, 0.37– 0.45 mm wide, apical portion conically tapered towards rather wide, dorsad-bent and rounded apex, which is in ventral and dorsal view conically attenuated and then constricted into thin, elongate-cylindric tip ( Figs 29, 31, 32 View FIGURES 19–32 ).
Variability. Elytral apex, which is mostly arcuate in middle, may possess an obtuse inner tooth; the elytral apex in female has variably developed external tooth. Only two, old and obviously badly treated specimens (MFNB) have faded setal vesture.
Distribution and biology ( Fig. 52 View FIGURE 52 ). Pogonostoma (P.) meridionale is phytogeographically separated from P. (P.) natsuae sp. nov. It inhabits the southernmost areas of Madagascar, and the label data of the above-listed specimens are in accordance with the distribution mentioned by Jeannel (1946), Horn (1934), Olsoufieff (1934) and Moravec (2007); only the localities confirmed by the examined specimens are marked in the simplified map of distribution ( Fig. 52 View FIGURE 52 ). P. (P.) meridionale may occur in different forest formations which in the wide area vary from the southernmost corner of the evergreen forest on the east coastal area near Tolagnaro, to the deciduous dried southern-inland and coastal forests and scrublands with prevailingly Euphorbia and Didiereaceae forests scattered west from Tolagnaro to Behara, Androy, Ambovombe, Ampasimpolaka and Imanombo. The type locality Fort Dauphin (now Tolagnaro) is not exactly specified and may represent different forest formations. We may speculate that the remnants of the dry and spiny formations originally covered larger areas northwards to Bekily and far westwards towards Toliara, also when the type and old historical specimens were caught. The male (CCJM), captured rather recently 24 km west of Fort Dauphin, might have been taken in either biotope. Many historical specimens come from Bekily, Behara, Ampasimpolaka in the Mandrar Basin and Andrahomana–Ambovombe. The localities Vall d’ Ambolo and Andrahomana are probably with remnants of evergreen eastern forest, while westwards localities around Ampasimpolaka are (or were) probably with prevailing scrublands. The label “Annanarivo of one male (SDEI) evidently means Madagascar (often labelled as such by historical insect dealers).
P. (P.) meridionale is now a rare species, known prevailingly from only old historical specimens. Apart from the above-mentioned recently caught male 24 km west of Tolagnaro, it was reported by Cassola & Violani (1992) from Berenty, which is a private natural reserve primarily focusing on the preservation and research of six species of lemurs. Hypothetically, this species has a chance to survive in the gallery forest of tamarind trees along the Man- draré River, and semi-arid spiny forest and scrubs in this small, but well-protected natural preserve.
Remarks. The shape of the aedeagus (previously illustrated by Moravec 2007), and its consistent difference from that in P. (P.) natsuae sp. nov. has been confirmed within this present revision.
Due to the dense elytral punctures, Rivalier (1970) placed P. (P.) meridionale in his “groupe meridionale ” together with diagnostically very different Pogonostoma (Transstenocera) sikorai W. Horn, 1934 , which inhabits evergreen forests of the northeastern and central-eastern areas of the island. Therefore, P. (P.) meridionale has been maintained in the P. (P.) elegans species-group (sensu Moravec 2007) as classified by Jeannel (1946).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Pogonostoma (Pogonostoma) meridionale Fleutiaux, 1899
| Moravec, Jiří, Razanajaonarivalona, Elisé Hugo & Hori, Michio 2020 |
Pogonostoma (Pogonostoma) meridionale:
| Moravec, J. 2007: 291 |
Pogonostoma (Pogonostoma) meridionale:
| Rivalier, E. 1970: 304 |
Pogonostoma (Pogonostoma) meridionale:
| Jeannel, R. 1946: 123 |
Pogonostoma meridionale
| Fleutiaux, E. 1899: 384 |