
Pogonostoma (Pogonostoma) natsuae, Moravec & Razanajaonarivalona & Hori, 2020
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4790.2.4 |
|
publication LSID |
lsid:zoobank.org:pub:6316903F-9894-40CF-A22C-8593637F1C9A |
|
persistent identifier |
https://treatment.plazi.org/id/038E87CB-D31A-CD01-FF21-F89D8306B66A |
|
treatment provided by |
Plazi |
|
scientific name |
Pogonostoma (Pogonostoma) natsuae |
| status |
sp. nov. |
Pogonostoma (Pogonostoma) natsuae View in CoL sp. nov.
( Figs 1–26 View FIGURES 1–9 View FIGURES 10–15 View FIGURES 16–18 View FIGURES 19–32 ; 52 View FIGURE 52 )
Type locality. Southwestern Madagascar (district of Atsimo-andrefana): isolated subhumid Analavelona Ankokony Forest (22°41´06´´S; 044°09´31´´E), 46 km northwest of Sakaraha GoogleMaps .
Type material. Holotype ♂ in OSAKA, labelled: “ Analavelona Ankokony Forest / Sakaraha (Atsimo-andrefa- na) / Region Madagascar / Dec. 1–4.2016 / Elisé Razanajaonarivalona leg.” [printed] . Allotype ♀ in OSAKA, with same locality label . Paratypes. 1 ♂ in MNHN , 1 ♂ in SDEI , 1 ♂, 1 ♀ in ZUAC , 3 ♂♂, 1 ♀ in CCJM , 2 ♂♂, 2 ♀♀ in CJVB , other 6 ♂♂, 6 ♀♀ in MHCW . The type specimens labelled: “ Holotype (“ Allotype or Paratype ” respectively) / Pogonostoma (s. str.) / natsuae sp. nov. / det. Jiří Moravec, / Elisé Hugo Razanajaonarivalona / & Michio Hori, 2019” [red, printed].
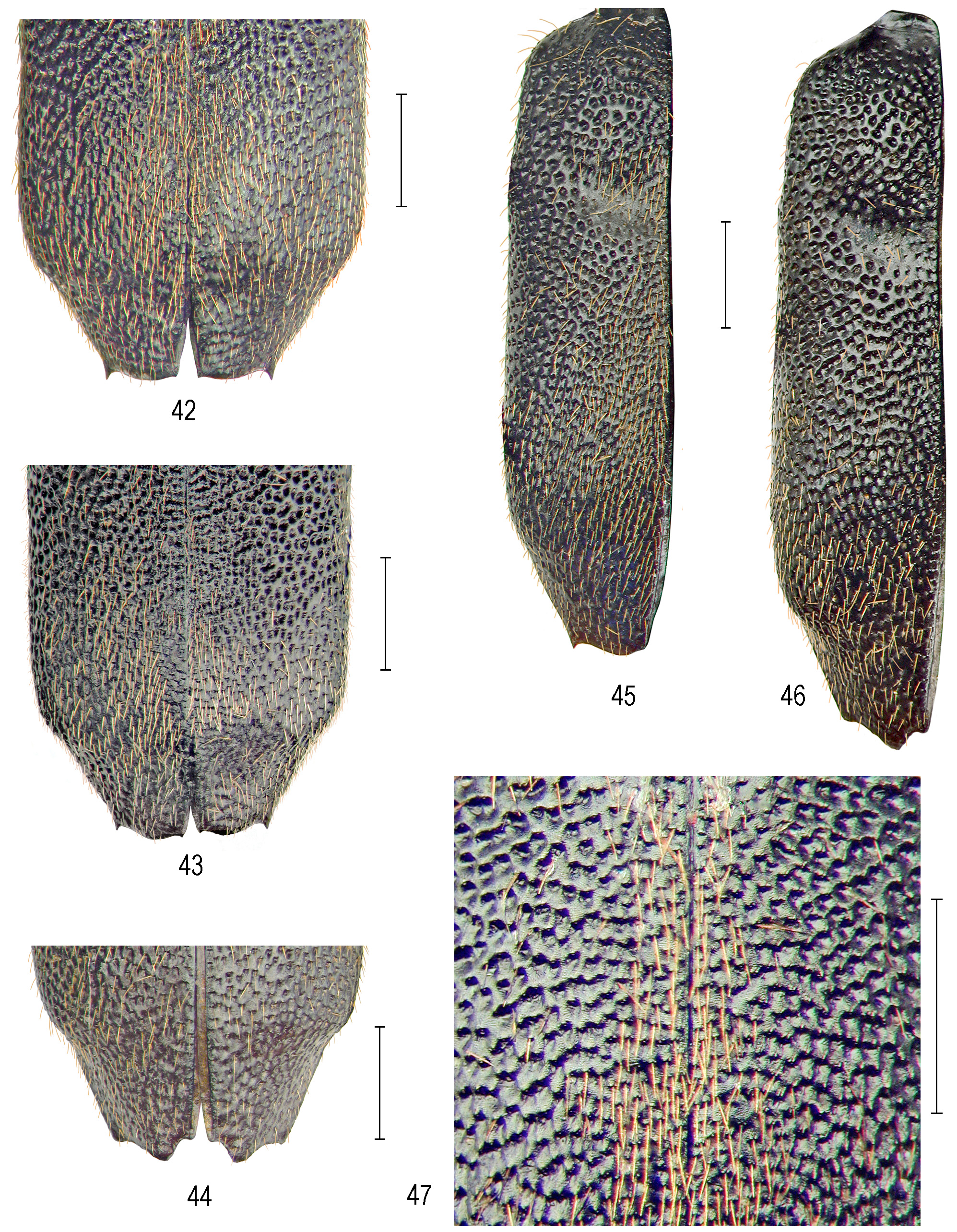
Differential diagnosis. Pogonostoma (P.) natsuae sp. nov. is considered here a species of the P. (P.) elegans species-group (sensu Moravec 2007). The new species is externally similar to P. (P.) meridionale Fleutiaux, 1899 , but can be distinguished by its more glossy elytra due to sparser and paler, blonde to straw-coloured ornamental setae, and elytral surface with generally larger and more regular punctures ( Figs 10–15 View FIGURES 10–15 ). Moreover, elytral apex possesses right-angled, rarely acute-angled inner tooth in both sexes ( Figs 10–14 View FIGURES 10–15 ). In contrast, the elytral punctation in P. (P.) meridionale is generally finer and more irregular, densest and with multiform intervals, particularly irregular along sutures, with more distinctly aciculate anterior margins on posterior elytral area, and the elytral apices are mostly sexually dimorphic in shape ( Figs 42–46 View FIGURES 42–47 ).
The lateral aspect of the aedeagus in both species appears somewhat similar, but the aedeagus of the new species is straighter ( Figs 19, 22 View FIGURES 19–32 ), while it is notably boomerang-bent in P. (P.) meridionale ( Figs 28, 30 View FIGURES 19–32 ). However, the major difference lies in the shape of the aedeagus apex in its ventral (and dorsal) aspect: while it is conically attenuated and nearly pointed in P. (P.) natsuae sp. nov. ( Figs 20, 21, 23, 24, 26 View FIGURES 19–32 ), it is constricted into thin, elongatecylindric tip in P. (P.) meridionale ( Figs 29, 31, 32 View FIGURES 19–32 ).
For differences from other relevant species of the (P.) elegans species-group see under P. (P.) meridionale below and in the Pogonostoma monograph ( Moravec 2007).
Description. Body 10.8–12.5 (HT 10.9, AT 12.3) mm long, 2.70–3.30 (HT 2.80, AT 3.10) mm wide, shiny black, setal vesture whitish-yellow to ochraceous or straw-coloured (the coloration changes depending on lightangle).
Head ( Figs 2–3 View FIGURES 1–9 ) notably narrower than body, width 2.30–2.60 mm; temples rather short, (slightly less than 2 times shorter than eyes).
Frons merging with clypeus in middle and not differentiated from vertex, flat or moderately convex; supraantennal keels consisting of elevated, blunt to acute anterior crest and usually less distinct posterior one; surface irregularly scabrous to alveolate.
Vertex moderately convex in middle with faint posteromedian impression, surface rather finely irregularly scabrous-alveolate, sculpture coarser on posterior area and passing to tubercles and short transverse-wavy rugae on occipital area; dorsal surface of head covered with erect, very short, whitish to ochre-yellow hairlike setae which are better obvious only in lateral view.
Genae finely or rather coarsely rugulose on posterior and juxtaorbital area, with several decumbent and erect, whitish, partly barely visible setae.
Clypeus black, rough-coriaceous, with dense, whitish or blonde, short setae.
Labrum primarily 6-setose, rarely 7-setose with 4–5 anterior and 2 lateral, ochre to ochre-brown setae, surface black, with moderate central convexity, rough-coriaceous, nearly glabrous or with scattered, barely obvious short microtrichia; lateral margins moderately arcuate towards indistinct lateral indentations, with rounded anterolateral margins (anterolateral teeth effaced), anterior margin irregularly dentate (with five to six small teeth); male labrum ( Figs 7–8 View FIGURES 1–9 ) 0.70–0.80 mm long, 1.20–1.35 mm wide, dentate anterior margin subtruncate to truncate; female labrum more prolonged anteriad, mostly semicircular, length 0.90–0.1.00 mm, width 1.40–1.50 mm.
Maxillae ( Fig. 6 View FIGURES 1–9 ): galea black, sometimes with brownish apex of basal (longest) galeomere; lacinia black with brownish, rarely reddish-brown faint tinge and with brownish or rusty setae; apical portion moderately clavate-dilated, 0.35–0.4 mm wide.
Palpi ( Fig. 2 View FIGURES 1–9 ) of usual length and shape, black, except for cinnamon-brown inner apical area of terminal palpomeres, covered with scattered, short whitish or greyish microtrichia, longest palpomeres of labial palpi with long, black-brown setae with mahogany to violet tinge.
Mandibles ( Figs 3–5 View FIGURES 1–9 ) with rather short terminal teeth, sexually dimorphic in shape, in male ( Fig. 3–4 View FIGURES 1–9 ) subsymmetrical (apart from the longer terminal tooth in right mandible as usual) with second tooth in both mandibles notably smaller than third tooth; female mandibles ( Fig. 5 View FIGURES 1–9 ) with inner teeth of right mandible almost of the same size.
Antennae black, in male as long as body or slightly longer, in female somewhat shorter; scape covered with rather dense whitish microsetae and with long, yellow-brown subanterior (easily abraded) setae; antennomeres 2–4 with rather numerous but scattered whitish microsetae, 5–11 with pubescence of greyish microtrichia.
Thorax: pronotum ( Figs 16–18 View FIGURES 16–18 ) elongate, 2.70–3.10 mm long, 1.85–2.10 mm wide; anterior lobe as wide as posterior lobe in female, in male often slightly narrower, its surface finely tuberculate and rugulose; pronotal disc in female mostly subglobose, in male with moderately convex, rarely subparallel lateral margins; in lateral view subgibbose (more distinctly in male, Fig. 17 View FIGURES 16–18 ); notopleural sutures invisible from above or very indistinctly at posterior pronotal angles in male; median line indistinct, merging with irregularly cristate-tuberculate surface sculpture consisting of tubercles commonly transversely connected into short, transverse-elongate crests, which are finer on anterior area and becoming larger on median area; posteromedian area adjacent to posterior lobe covered with fine and shallow, more transverse-continuous stria-like rugae; whole discal surface covered with scattered, short, decumbent or erect, whitish to ochre-yellow or straw-yellow hairlike setae which are barely obvious in dorsal view, but well recognizable in lateral view ( Fig. 17 View FIGURES 16–18 ); posterior lobe shiny and smooth except for few indistinct shallow wrinkles in middle, entirely glabrous; proepisterna large, polished and smooth, finely and shortly parallel-wrinkled on juxtanotopleural area, nearly glabrous, except for only occasional hairlike setae; mesepisterna large, polished, with only occasional setae, mesepimeron deeply impressed; metepisterna notably long, deeply impressed, smooth and glabrous; prosternum smooth, except for very fine transverse wrinkles, rather densely covered with long and erect, whitish hairlike setae arising from barely visible setigerous micropunctures; mesosternum sparsely punctate-setose with shorter whitish setae; metasternum almost smooth, indistinctly sparsely whitish setose, glabrous in middle.
Elytra ( Figs 10–15 View FIGURES 10–15 ) elongate, 6.00– 7.30 mm long; humeri obliquely arcuate; lateral margins almost parallel ex- cept for slightly arched bulge in anterior third of the margin in male, and only indistinctly enlarged towards rounded lateral anteapical angles; elytral apices almost uniform in both sexes, with faint sutural emargination towards indistinct sutural spine, and with right-angled, rarely acute-angled inner tooth between external tooth which is thornlike in male, while blunter and right-angled in female; elytral surface convex with moderate basodiscal convexity delineated by deep discal impression, punctate throughout (except for effaced narrow basal and basohumeral area); punctures large and deep on two elytral thirds, large and almost isolated on lateral areas, deepest, large and irregularly anastomosing by lowered lateral intervals on basodiscal convexity, reduced or partly (never entirely) effaced within discal impression, and again large but only occasionally anastomosing on elytral disc except for much finer and irregular sculpture on limited area along sutures; punctures become smaller, more spaced and with aciculate anterior margins of intervals on posterior declivity and whole anteapical area, but larger and irregular on apices; surface of intervals shiny; setal vesture consisting of nearly erect, copious, moderately long, pale to straw-yellow ornamental setae which are mixed with very sparse, long and erect, whitish hairlike sensory setae, which are very long and numerous on lateral portion of humeri.
Abdomen. Ventrites black, sometimes with very faint diffusing bluish lustre, covered with short, dense, but scattered and mostly appressed white setae.
Legs black; coxae rather densely whitish-setose; profemora covered with rather dense medium-long whitish setae; mesofemora with dense whitish setae on their dorsal surface, much sparser setae on ventral area; metafemora with sparser, ochre and brownish setae; pro- and mesotibiae with whitish setae except for dense, ochre-ferrugineous setae on apices of tibiae; tarsi dark reddish-brown, their surface covered with whitish setae (and as in all species, first three protarsomeres in male dilated and with dense setose pad); claws rusty-brown.
Aedeagus 2.50–2.60 mm long, 0.40 mm wide, in its left lateral aspect ( Figs 19, 22, 25 View FIGURES 19–32 ) rather distinctly but gradually bent in middle, apical portion conically tapered towards rather narrow and small, dorsad-bent, rounded apex, which is in its ventral (and dorsal) aspect conically attenuated and nearly pointed ( Figs 20, 21, 23, 24, 26 View FIGURES 19–32 ).
Distribution and biology ( Fig. 52 View FIGURE 52 ). The type locality in the Analavelona Massif 46 km northwest of Sakaraha is characterized as a subhumid forest on basalt at an elevation of 700–1325 m. The adults of the new species were caught by the second author on trunks of high but not very large trees at a higher altitude. This uniquely isolated forest, surrounded by dry succulent forest formations at lower elevations, is maintained owing to a condensation of moisture from ascending air. The unique system of the Analavelona Massif encompasses 77 tree and plant families with high level of endemism ( Randrianarivony 2015). As specified by Goodman et al. (2018), the subhumid climate is associated with a local orographic effect on the upper portions of the massif with an average annual rainfall of around 856 mm, 93% usually falling between November and April. The basaltic substrate probably contributes to the persistence of a particularly moist type of forest of the region which justified the massif being the western most extension of the phytogeographical zone known as the Central Domain. Given its multi-layered composition with absence of Uapaca bojeri Baill trees ( Phyllanthaceae ), this forest is not sclerophylous woodland, despite its geographical placement. In fact, it has characteristics of medium altitude moist evergreen forest, as of the 75 recognized tree species only four are deciduous. For other details and the list of tree species see Goodman et. al. (2018). Fortu- nately, the main parts of the nice and now newly protected forest have remained almost untouched, being a sacred place for the Malagasy local Bara people, who believe that it is the home of the spirits of their ancestors.
Two other tiger beetle species were caught at the same time in the type locality of the new species: Pogonostoma (Microstenocera) zombitsynense Moravec & Vybíral, 2010 and Physodeutera (Diarrhiza) tricolorata (W. Horn, 1934) .
Etymology. This species is cordially dedicated to Madame Natsu Hori, the recently deceased mother of the third author.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |