
Haplochromis rex, Vranken & Steenberge & Heylen & Decru & Snoeks, 2022
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.815.1749 |
|
publication LSID |
lsid:zoobank.org:pub:6AD0082E-7349-48DE-AFCA-1EE0BFBB3887 |
|
DOI |
https://doi.org/10.5281/zenodo.6484175 |
|
persistent identifier |
https://treatment.plazi.org/id/EF8B7189-BCA4-4A72-8387-498689D76CEB |
|
taxon LSID |
lsid:zoobank.org:act:EF8B7189-BCA4-4A72-8387-498689D76CEB |
|
treatment provided by |
Felipe |
|
scientific name |
Haplochromis rex |
| status |
sp. nov. |
Haplochromis rex sp. nov.
urn:lsid:zoobank.org:act:EF8B7189-BCA4-4A72-8387-498689D76CEB
Figs 1–2 View Fig View Fig , 11–13 View Fig View Fig View Fig ; Table 1 View Table 1
Differential diagnosis
Species with a piscivorous morphology; head narrow [HW 36.8–41.6 (mean 39.2) % HL]; cheek deep [ChD 27.6–33.5 (31.1) % HL]; eye small [ED 22.2–28.3 (24.6) % HL]; outer oral teeth few and large [UOT 24–36 (median 29)]; dominant males cream-coloured with an orange operculum and a light blue snout.
Amongst piscivorous species from the Lake Edward system, H. rex sp. nov. differs from H. latifrons sp. nov. and H. mentatus by the combination of a shorter caudal peduncle [CPL 13.5–16.2 (14.8) vs 15.7–18.0 (16.6–17.0) % SL] and a narrower interorbital area [IOW 44.9–52.7 (48.9) vs 51.3–63.3 (55.5–60.0) % HW]; further from H. latifrons sp. nov. by absence vs presence of a well-defined midlateral band; further from H. mentatus by dominant males cream-coloured with an orange operculum vs yellow-green with a red anterior part of flank.
It differs from H. simba sp. nov. by the combination of a larger number of longitudinal line scales (LongL 34–38 vs 32–33), a larger number of scales between first anal-fin spine and upper lateral line (ULL-A 12–16 vs 9–11), absent or weakly developed vs strongly developed mental prominence, and dominant males cream-coloured with an orange operculum and a light blue snout vs yellow with an orange anterior part of flank and a yellow snout.
It differs from H. glaucus sp. nov. by the combination of a narrower interorbital area [IOW 44.9–52.7 (48.9) vs 50.9–57.1 (53.8) % HW], a steeper snout (40–50° vs 30–40°), rounded vs acute oral jaws in dorsal view, and dominant males cream-coloured with an orange operculum and a light blue snout vs uniformly light blue with a dusky snout.
It differs from H. aquila sp. nov. by the combination of a deeper lacrimal [LaD 18.9–22.5 (20.8) vs 17.0–19.1 (18.3) % HL], smaller eye [ED 22.2–28.3 (24.6) vs 30.0–31.5 (30.6) % HL], and dominant males cream-coloured with an orange operculum vs light grey with a black head.
It differs from H. kimondo sp. nov., H. falcatus sp. nov., H. curvidens sp. nov., H. pardus sp. nov., H. quasimodo sp. nov., and H. squamipinnis by the combination of stout vs slim oral jaws, large vs small outer oral teeth, and a smaller number of outer upper jaw teeth [UOT 24–36 (29) vs 39–79 (45–58)].
It further differs from H. kimondo sp. nov., H. curvidens sp. nov., and H. quasimodo sp. nov. by a narrower head [HW 36.8–41.6 (39.2) vs 42.0–48.1 (43.4–45.3) % HL]; further from H. kimondo sp. nov. and H. quasimodo sp. nov. by dominant males cream-coloured with an orange operculum vs grey dorsally and yellow or blue-black ventrally.
It further differs from H. falcatus sp. nov., H. pardus sp. nov., and H. squamipinnis by a larger number of longitudinal line scales (LongL 34–38 vs 29–33, rarely 34) and dominant males cream-coloured with an orange operculum vs uniformly olive-green with an orange-red anterior part of flank, speckled to uniformly black, or uniformly slate blue, respectively.
Etymology
Specific name from the Latin ‘ rex ’ for ‘king’ (one that holds a preeminent position); referring to very small eyes, deep cheeks, and strong jaws set with large and acute teeth indicating this piscivore has most specialised morphology among all piscivores from the Lake Edward system to hunt on large prey ( Barel et al. 1977).
Material examined
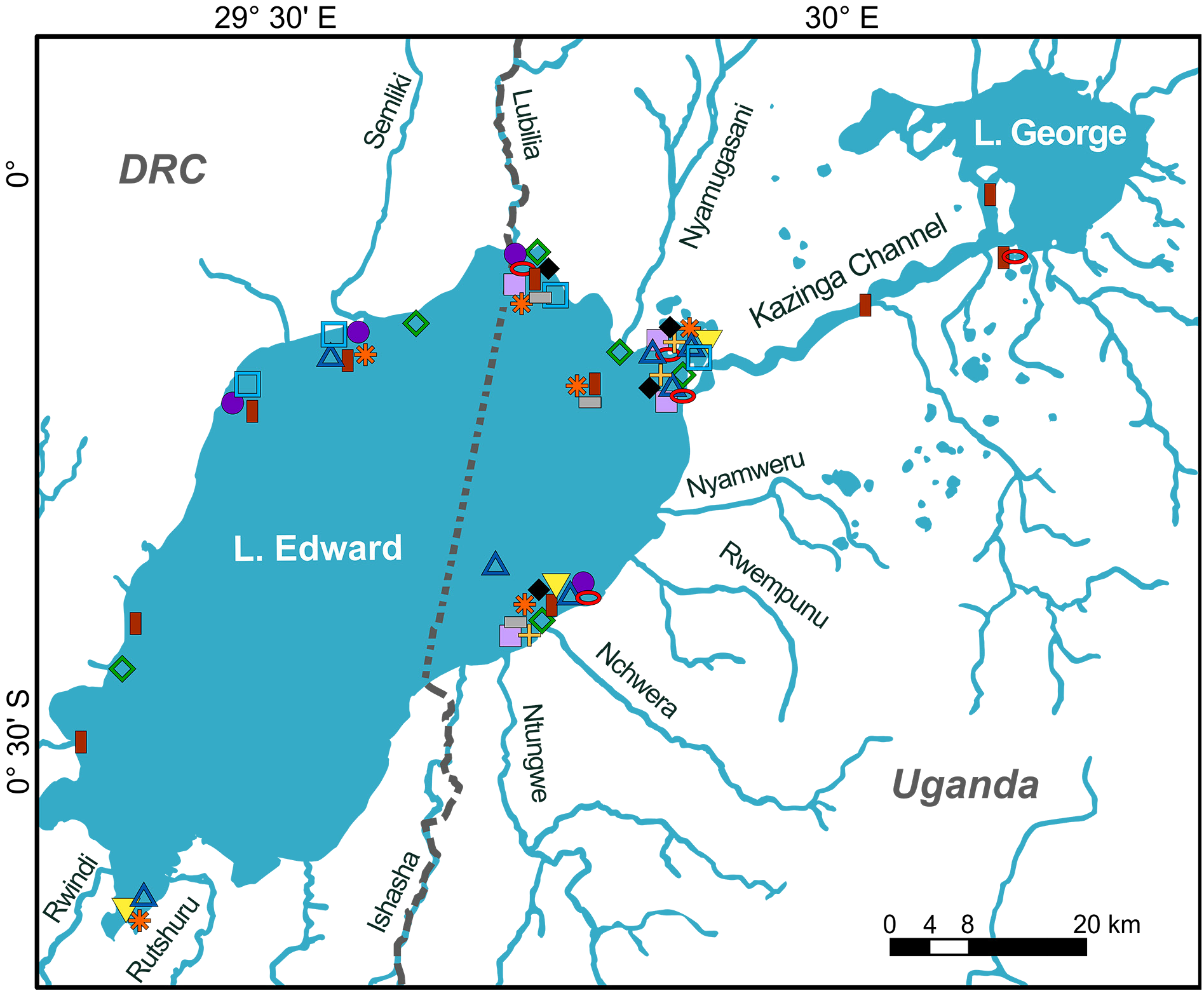
Holotype UGANDA • ♂, 148.5 mm SL; Lake Edward; 0°24′16.0″ S, 29°46′24.8″ E; 24 Jan. 2018; HIPE3 exped. leg.; bought at Rwenshama landing site; RMCA 2018.008.P.0345 . GoogleMaps
Paratypes DEMOCRATIC REPUBLIC OF THE CONGO • 1 ♂, 131.5 mm SL; “Lac Edouard: Vitshumbi (contre le pier)” [Lake Edward: Vitshumbi (against the pier)]; 0°40′50.6″ S, 29°23′22.6″ E (inferred); 26 Mar. 1953; KEA exped. leg.; IRSNB 13474 View Materials GoogleMaps • 1 ♂, 154.2 mm SL; “Lac Edouard: 2–3 km ± 500 m au large à l’Ouest de Kiavinionge” [Lake Edward: 2–3 km ± 500 m offshore west of Kiavinionge ]; 0°11′39″ S, 29°32′31″ E (inferred); 1 Jun. 1953; KEA exped. leg.; IRSNB 13480 View Materials GoogleMaps • 1 ♀, 154.8 mm SL; “Lac Edouard: 2–3 km à l’Ouest de Kiavinionge” [Lake Edward: 2–3 km west of Kiavinionge ]; 0°11′39″ S, 29°32′31″ E (inferred); 1 Jun. 1953; KEA exped. leg.; IRSNB 13485 View Materials GoogleMaps .
UGANDA – Lake Edward • 1 ♂, 114.4 mm SL; 0°12′00.0″ S, 29°47′38.4″ E; 23 Oct. 2016; HIPE1 exped. leg.; deep catch, open water ± 20 m deep; RMCA 2016.035.P.0220 GoogleMaps • 1 ♂, 154.5 mm SL; 0°08′51.9″ S, 29°52′59.6″ E; 24 Oct. 2016; HIPE1 exped. leg.; bought at Katwe landing site; RMCA 2016.035.P.0221 GoogleMaps • 1 ♂, 114.7 mm SL; Rwenshama , rocky shore; 0°24′05.7″ S, 29°46′35.1″ E; 26 Mar. 2017; HIPE2 exped. leg.; RMCA 2017.006.P.0352 GoogleMaps • 1 ♂, 2 ♀♀, 101.0–140.0 mm SL; Kayanja offshore; 0°05′34.8″ S, 29°45′28.8″ E; 31 Mar. 2017; HIPE2 exped. leg.; RMCA 2017.006.P.0353 to 0355 GoogleMaps • 1 ♀, 80.5 mm SL; Kayanja offshore; 0°05′31.2″ S, 29°45′30.3″ E; 20 Jan. 2018; HIPE3 exped. leg.; RMCA 2018.008.P.0342 GoogleMaps • 1 ♂, 1 ♀, 134.6, 147.7 mm SL; 0°24′16.0″ S, 29°46′24.8″ E; 24 Jan. 2018; HIPE3 exped. leg.; bought at Rwenshama landing site; RMCA 2018.008.P.0343 to 0344 GoogleMaps • 1 ♀, 84.2 mm SL; Rwenshama , offshore; 0°24′14.4″ S, 29°45′57.0″ E; 24 Jan. 2018; HIPE3 exped. leg.; RMCA 2018.008.P.0346 GoogleMaps • 1 ♂, 128.3 mm SL; Kagoro fishing ground; 0°12′50.1″ S, 29°49′19.7″ E; 4 Feb. 2018; HIPE3 exped. leg.; open water; RMCA 2018.008.P.0347 GoogleMaps • 1 ♀, 134.2 mm SL; Kayanja offshore; 0°05′34.8″ S, 29°45′28.8″ E; 21 Mar. 2019; HIPE4 exped. leg.; RMCA 2019.002.P.0014 GoogleMaps .
Description
Based on 16 specimens (80.5–154.8 mm SL); body shallow ( Table 1 View Table 1 ) and oval to rectangular ( Fig. 11 View Fig ). Head long, very narrow, and with a (weakly) convex dorsal outline; cheek deep; lacrimal very deep; eye very small and very high on head; interorbital area very narrow. Snout long, rounded in dorsal view, blunt, and slopes at 40–50°; premaxillary pedicel long and strongly prominent. Jaws isognathous to weakly prognathous, long, very narrow, stout, and rounded in dorsal view; jaws expand slightly laterally, hereby often broader than snout. Gape large and slopes gently at 15–25°; maxilla extends to vertical through anterior margin of orbit. Lower jaw relatively deep and with a straight ventral outline in lateral view, mental prominence absent or weakly developed, and lower jaw side with an inclination of 30–35° to horizontal in anterior view. Upper jaw strongly expanded anteriorly. Lips and oral mucosa large. Neurocranium shallow, ethmo-vomerine block decurved, preorbital region very shallow (18–20% NL), orbital region shallow (26–29% NL), and supraoccipital crest shallow and wedge-shaped ( Fig. 12b View Fig ).
Outer oral teeth few, unicuspid, and large. Necks very stout, conical, and straight; crowns weakly recurved, rarely straight, and acutely pointed. Dental arcades rounded. Outer teeth widely and regularly set with neck-distances of 1–2 neck-widths. No enlarged teeth posterior in upper jaw. Inner teeth straight to recurved, unicuspid, and relatively small in small specimens (<125 mm SL), and large in large specimens (> 125 mm SL). Tooth bands very slender crescent-shaped with 1–3 rows of inner teeth, and narrow posteriorly until only outer row remains past ⅔ length of tooth band in upper jaw, past ½ length of tooth band in lower jaw. Inner teeth widely and regularly set on 1–2 outer neck-widths from outer row; implantation erect in first row and recumbent in subsequent rows; size decreases slightly buccally and posteriorly.
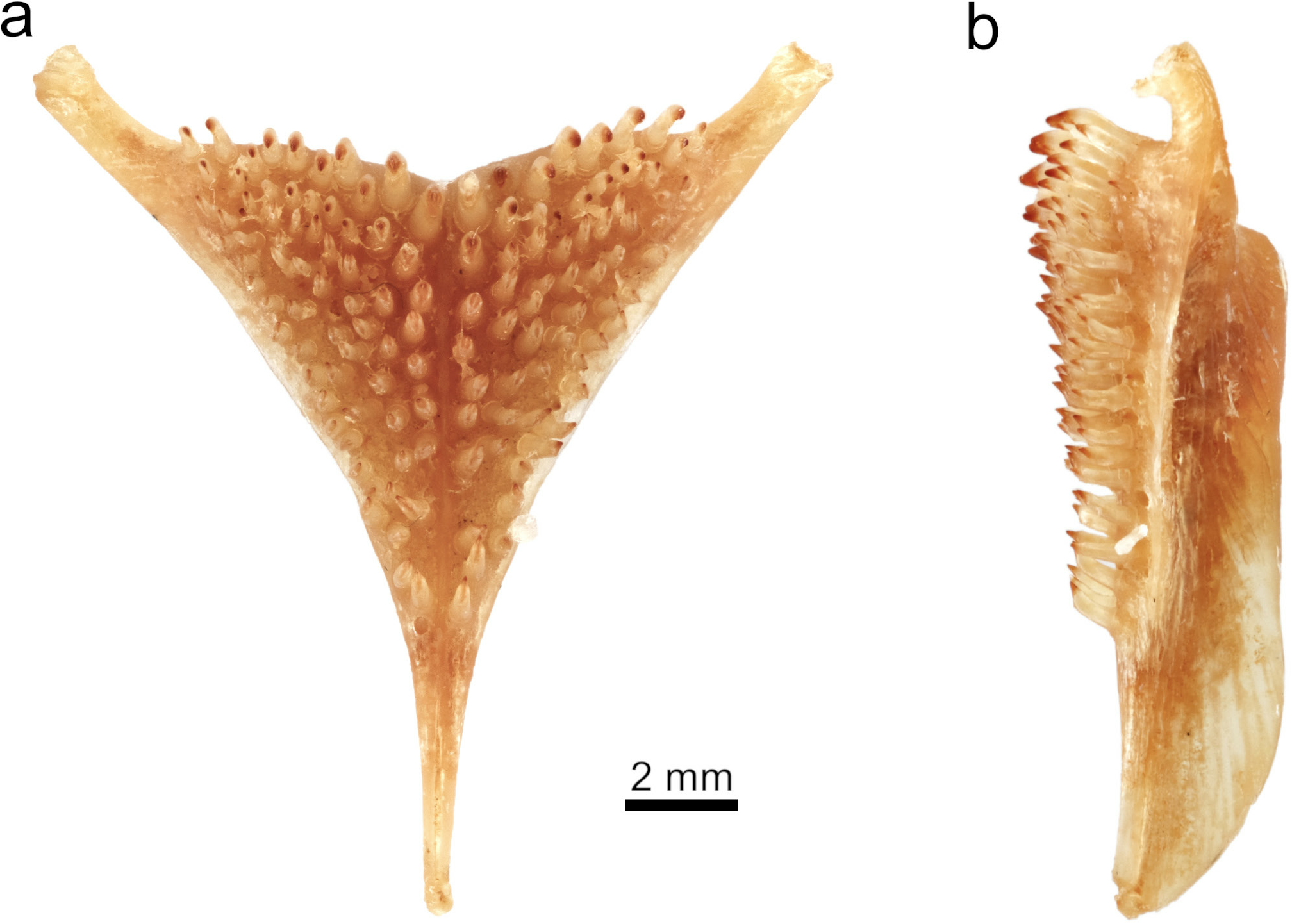
Lower pharyngeal bone average in length, narrow, slim, and shallow over whole length ( Fig. 13 View Fig ). Pharyngeal teeth relatively large and slender; major cusps acutely pointed; cusp gaps concave; minor cusps and cusp protuberances mostly absent. Teeth in two median longitudinal rows equal in size and form to lateral teeth, 11 in each row. posterior transverse row with 15–16 teeth, implanted recumbently with a lateral inclination; major cusps weakly recurved, bluntly pointed, and laterally compressed; minor cusps mostly absent.
All scales relatively small, many scale rows between anal fin and upper lateral line (12–16), infraorbital scales on cheek (4–7), and scales around caudal peduncle (18–20); many scales in longitudinal line (34–38); chest scales small; transition to larger flank scales gradual. Minute scales on proximal half of caudal fin.
Caudal fin emarginate; dorsal and anal fins reach to between verticals through two scales anterior to and one scale posterior to caudal-fin base. Pectoral fin reaches to between anal opening and three scales anterior to this point; pelvic fin reaches to anal opening in females, to first anal-fin spine in males; first branched pelvic-fin ray not elongated.
Ceratobranchial gill rakers in outer row of first gill arch short, stout, and simple; posteriormost rakers anvil-shaped to weakly trifid. Epibranchial gill rakers relatively slender and simple.
Colouration in life
Dominant males: body cream-coloured with faint orange sheen; dorsum greyish; belly and chest black; caudal peduncle with blue sheen ( Fig. 12c View Fig ). Operculum, dorsal part of head, and interorbital area orange; snout, lower jaw, and lips light blue; cheek white and speckled black; eye with (dark) grey outer ring and silver inner ring. Nostril, interorbital, supraorbital, and lacrimal stripes faint; mental blotch present. Pectoral and dorsal fin hyaline; dorsal fin with a dusky base, a dusky and maculated posterior part, and orange lappets, except for black anteriormost lappets. Pelvic fin black; anal and caudal fins orange-red and with dusky bases; anal fin with dusky posterior part and 1–3 small orange egg-spots with dusky rings; caudal fin dorsally maculated orange.
Females and juveniles: body cream-coloured with yellow sheen; dorsum and dorsal part of head greyish; belly, chest, operculum, cheek, and lower jaw white; snout dusky and faint light blue; eye with (dark) grey outer ring and silver inner ring ( Fig. 12d View Fig ). Nostril, interorbital, supraorbital, and lacrimal stripes slender and well-defined; mental blotch present. Pectoral, pelvic, anal, and caudal fins yellowish; anal fin with 2 spots resembling egg-spots; caudal fin with dusky base and dorsal part; dorsal fin dusky and with black lappets.
Preserved colouration
Body, belly, operculum, and cheek yellowish; dorsum brown; chest yellowish in females, black in males ( Fig. 12a View Fig ). Flank rarely with 7–8 very faint narrow vertical stripes. Snout dusky; in males, lower jaw black. In females, all head stripes faint and lacrimal stripe reduced to blotch below eye; in males, head stripes as described here. Nostril and interorbital stripes narrow, well-defined, and horizontally inclined in lateral view, while positioned anterior of nostrils and orbits, respectively, in dorsal view; lacrimal stripe distinct, broad (may cover whole lacrimal), and vaguely delineated; vertical preopercular stripe welldefined; supraorbital stripe present; nape band and mental blotch faint. Pectoral and anal fins yellowish; anal fin with dusky base and posterior margin and 2 small egg-spots in males; pelvic fin yellowish with a black first ray in females, black in males. Dorsal and caudal fins dusky and with yellowish posterior parts; dorsal fin with black lappets and maculated posterior parts.
Distribution and ecology
Only known from Lake Edward, found over sandy substrates. Based on its morphology, most probably a piscivorous species.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |