
Hipposideros khaokhouayensis, Guillén-Servent & Francis, 2006
|
publication ID |
https://doi.org/ 10.3161/1733-5329(2006)8[39:ansobo]2.0.co;2 |
|
persistent identifier |
https://treatment.plazi.org/id/038F87BA-2543-FFF5-FF49-FD2D1EE7FE83 |
|
treatment provided by |
Carolina |
|
scientific name |
Hipposideros khaokhouayensis |
| status |
sp. nov. |
Hipposideros khaokhouayensis View in CoL sp. nov.
( Figs. 2–5 View FIG View FIG View Fig View FIG , Table 1 View TABLE )
Etymology
The name is derived from the Phou Khao Khouay NBCA where the type series was collected. This name was selected to bring attention to the potential importance ofthe National Biodiversity Conservation Areas network for preserving natural areas in Lao PDR, as they contain highly distinc- tive ecosystems, hosting many endemic species such as this one. Unfortunately, even within the NBCA, habitats are at risk: at the time of publication of this paper, the locations where the species was captured within Phou Khao Khouay NBCA were be- ing affected by deforestation, road work and building associated with the develop- ment of the large Tak Leuk dam project.
Holotype
Royal Ontario Museum, ROM 116790 (Field No. CMF980203-29), adult ♀, body in ethanol, skull extracted, collected by Charles M. Francis (CMF) on 3rd February 1998 as part of a WCS survey.
Type locality
Tak Leuk waterfall, along Nam (riv- er) Leuk, in Phou Khao Khouay National
Biodiversity Conservation Area, Saisombun Special Zone, Central Lao PDR, 18°23’N, 103°04’E, 180 m elevation.
Paratypes
ROM 116791 (Field No. CMF 980203- 26), adult ♂; ROM 110671 (Field No. CMF 980203-27), adult ♂; both bodies in ethanol, skulls extracted, collected by CMF on same date and locality as the holotype.
Estación Biológica de Do ñ ana, Sevilla, Spain (EBD) 23514 (Field No. AGS 970604-01), adult ♀, body in ethanol, skull extracted, collected by Antonio Guillén- Servent (AGS) on 4th of June 1997, at a logging road in Dan Phorkeng area near Dam on Nam (river) Leuk, 18°26’N, 102°57’E, 400 m elevation; EBD 23518 (Field No. AGS 970605-12), adult ♀, body in ethanol, skull extracted, collected by AGS on 5th of June 1997 in a place named Danpapor, located near Ban (village) Hoileuk, 18°34’N, 102°59’E, 260 m eleva- tion. Both localities are within the Phou Khao Khouay NBCA, near the site where the holotype was collected.
Instituto de Ecología, A. C., Xalapa, Veracruz, México (IEX) M0106, ROM 116792, IEX-M0107, ROM 116793 (Field Nos. AGS 980511-03, 980511-13, 980511- 14, 980511-15), 2 ♂♂ and 2 ♀♀ adult bats, respectively, bodies in ethanol, skulls ex- tracted, collected by AGS on 11 May 1998 near Ban Nampe, about 11 km NE of Vang Vieng, Vang Vieng District, Vientiane Prov- ince, Central Lao PDR, 18°55’N, 102°25’E, 240 m elevation.
Measurements
First number after variable name corre- sponds to the holotype. Minimum, maximum (separated by a dash), and mean of the type series are indicated between paren- theses (sample size is always eight except when stated otherwise as n = sample size).
All measurements (see also Table 1 View TABLE ) in mm, except body masses, which are in grams.
Body. Head and body 49.26 (44.13– 49.98, 47.16); tail 35.05 (34.95–37.51, 36.22); hindfoot 7.45 (7.10–8.27, 7.68); ear 25.24 (24.36–25.24, 24.79); forearm 47.02 (45.54–48.55, 47.12); metacarpal III 34.65 (32.21–34.65, 33.79); metacarpal IV 35.61 (35.05–36.03, 35.44); metacarpal V 33.57 (32.87–34.90, 33.92); tibia 20.16 (20.16– 22.07, 20.94); calcar 12.24 (11.35–12.24, 11.83); body mass 8.8 g (7.7–9.6, 8.7).
Width of noseleaves. Posterior 7.26 (6.37–7.86, 6.95); intermediate 5.33 (4.90– 5.44, 5.17); horseshoe 6.36 (5.23–6.72, 6.22); internarial septum 1.69 (1.5–1.81, 1.65). Skull. Greatest length (to incisors) 19.66 (19.18–20.04, 19.65); ibid. to C 1 at crown 19.26 (18.89–19.81, 19.30); ibid. to C 1 at alveola 18.28 (18.01–18.57, 18.23); condylobasal length (to incisors) 17.71 (17.05–18.27, 17.68); condylocanine length (to front of canine) 17.05 (16.02–17.56, 17.03); ibid. to alveola of canine 16.28 (15.95–16.75, 16.31); mastoid width 10.04 (9.86–10.37, 10.06); breadth of braincase 9.97 (9.84–10.41, 10.09); zygomatic width 9.85 (9.42 – 9.93, 9.67); width across ante- orbital foramen 5.23 (5.01–5.55, 5.21); width across rostral inflations 5.55 (5.21–5.57, 5.43); C 1 –C 1 breadth 4.05 (3.95–4.31, 4.11); M 3 –M 3 breadth 6.47 (6.12–6.47, 6.27); C–M 3 at crowns 6.74 (6.41–6.74, 6.62); ibid. C–M 3 at alveoli 6.13 (5.59–6.13, 5.92); interorbital con- striction 2.89 (2.73–3.03, 2.86); bulla length 3.44 (3.32–3.62, 3.41); basioccipital width between cochleae 1.33 (1.33–1.51, 1.41); width of the nasal channel frontal opening 2.57 (2.33–2.57, 2.47); palatal length 2.56 (2.39–2.86, 2.65); mandible length from condylus 11.82 (11.63–12.24, 11.87); ibid. from angular process 11.49 (11.27–11.85, 11.54, n = 7); C–M 3 length at
crowns 7.19 (6.76–7.25, 7.08); C–M 3 length at alveoli 7.10 (6.67–7.20, 6.95).
Diagnosis
Abat in the H. bicolor subgroup ( Hill, 1963) similar in size and general aspect to the sympatric H. rotalis , also with large ears, but differing in smaller and narrower noseleaf without supplementary leaflets; smaller, oval internarial disc only moderate- ly developed; smaller rostral inflations on skull ( Fig. 3 View FIG ); and higher frequency echolo- cation calls (constant frequency component 87–91 kHz). Externally similar to H. orbi- culus from southern Peninsular Malaysia and Sumatra, but differing in shorter tail; rostral chambers with different configura- tion and much less bone density and less walls; zygomatic arch with dorsal process very low and inconspicuous; bullae larger and bassioccipital much narrower between cochleae; second lower incisors with cin- gulid heel; upper canines without posterior cingulid cusp; and higher frequency echolo- cation calls.
Description ( Figs. 2 View FIG and 3 View FIG )
This is a moderate-sized Hipposideros ofthe H. bicolor subgroup ( Hill, 1963). Dorsal pelage is long and brown, with hair light brown to the tips, dark brown on the distal third of their length, and white on the basal two-thirds. Ventral pelage is pale with a brownish tinge, with hair very light brown to the tip and white to the base. Pelage is lighter on the front between the ears than on the back, and dark around the eyes. Aband of dark pelage runs down along the sides of the neck, fading progressively to the front. Ears brown in color, enlarged, long and broad, with tips bluntly pointed. Awell de- fined internal fold runs next and parallel to the inner border of the ear. A dense line of hair runs parallel to this fold on its outer side, and another one next to the inner bor- der of the ear. Noseleaf well developed, but
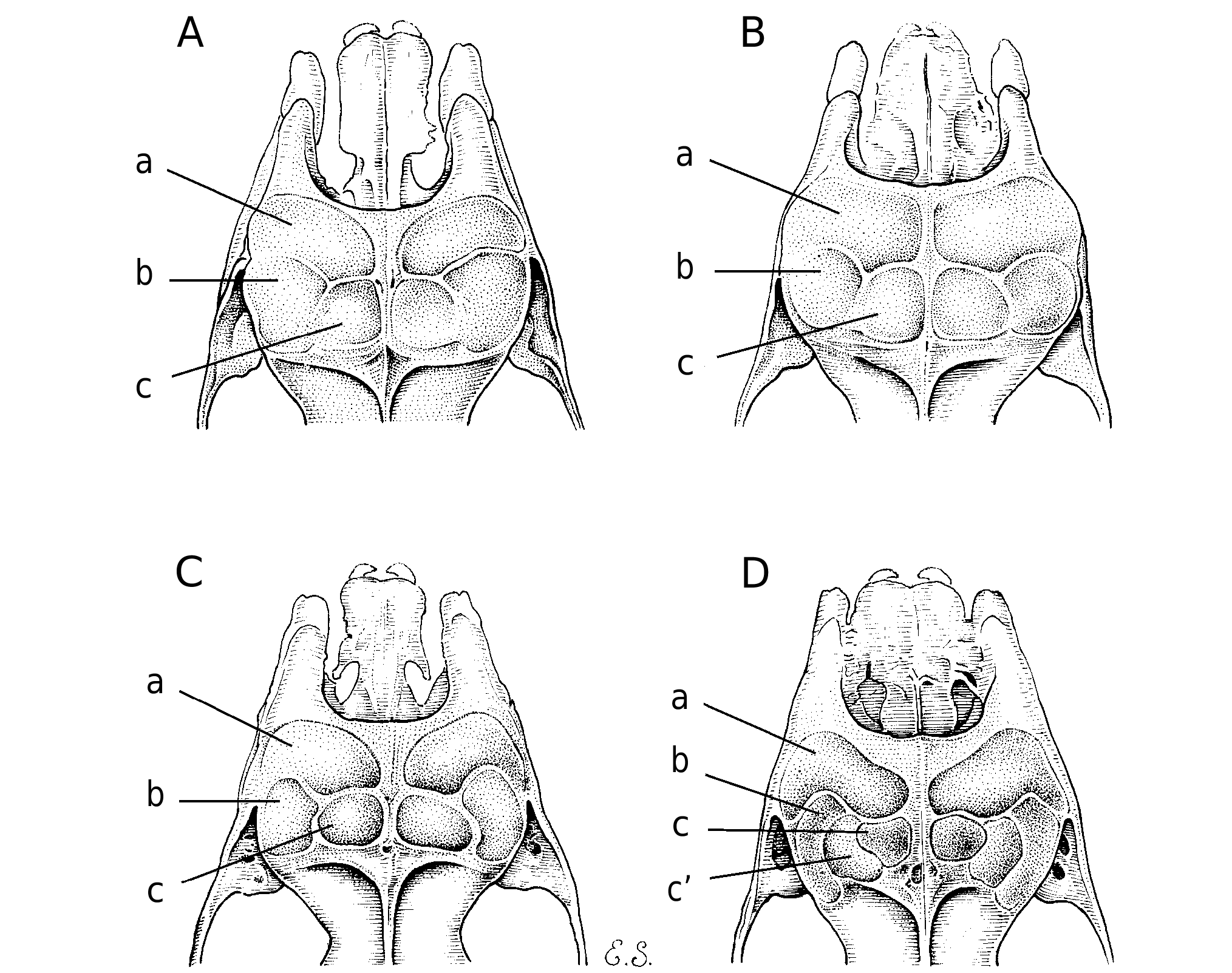
relatively simple in structure. No supplementary leaflets beneath horseshoe, but there is a longitudinal swelling behind and parallel to its outer margin, which may rep- resent an incipient leaflet. The anterior leaf has a narrow median emargination on its frontal margin. The internarial septum is expanded into a small oval disc between the nostrils. There are two pairs of papillae, with a single vibrissa each, on the intermediate leaf. Posterior leaf supported with three septa that define four pouches extending frontally to the upper margin of the in- termediate leaf, which are partially covered by the high upper margin of the posterior leaf. The bats have a conspicuous frontal glandular sac containing a tuft of hairs shorter, stiffer, and darker than surrounding fur. This sac is a little less developed in females. Tail moderately long, with tip not protruding from the uropatagium.
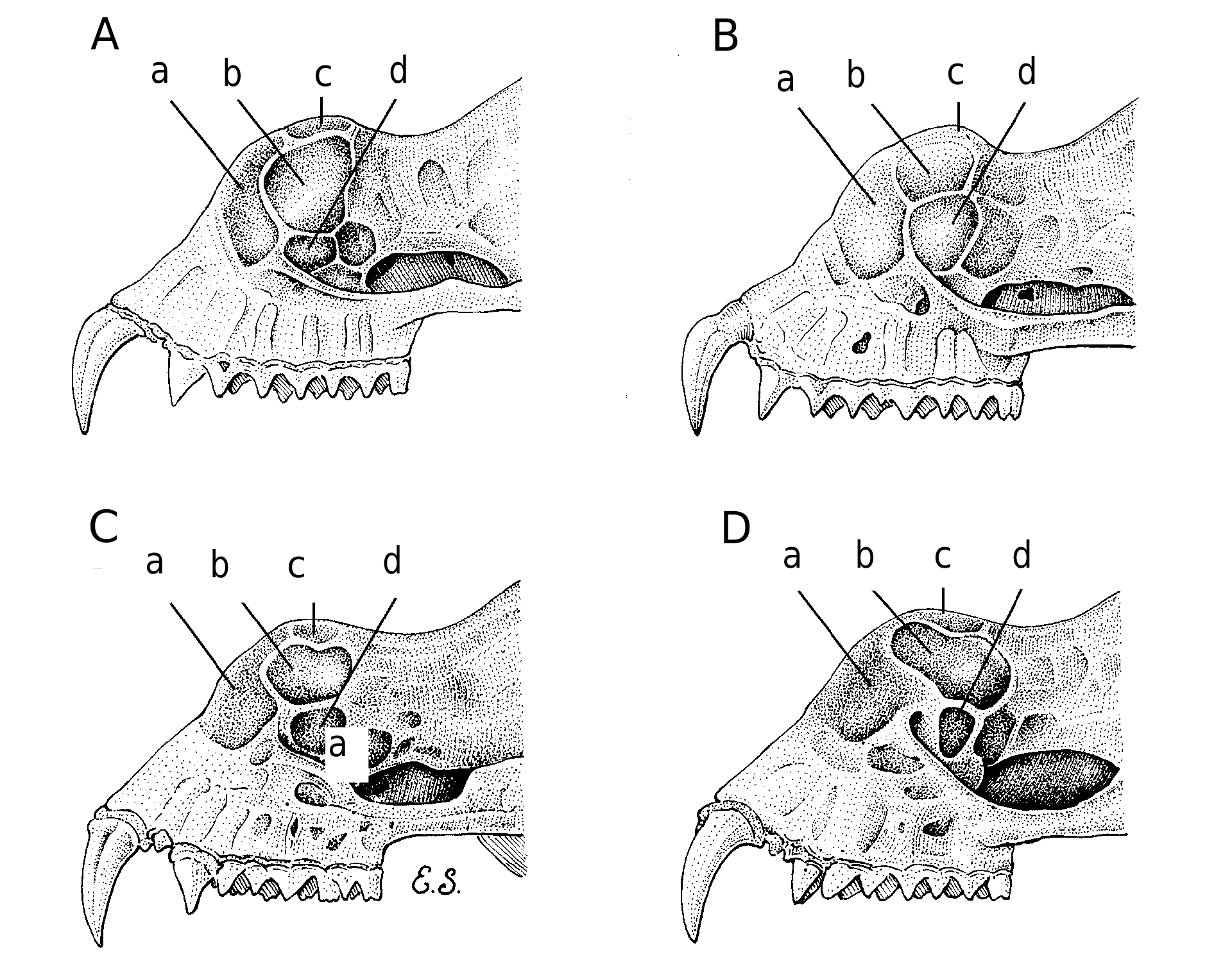
Skull ( Fig. 4 View Fig ): Anteorbital foramen large and elongated, with bone bridge from lateral rostral inflations to zygomatic arch very thin. Rostral inflations developed, but much less than in H. rotalis . Anterior rostral chamber extends laterally down to lower margin of anteorbital foramen. Posterior median rostral chamber approximately square in shape and the same length as the anterior rostral chamber along the sagital suture. Postero-lateral rostral chamber much enlarged, up to twice that of the pos- terior median chamber, and projecting down into the lachrymal region above the anterior orbita, leaving a much reduced lachrymal chamber, about one fifth the size of the postero-lateral one ( Figs. 5 View FIG and 6 View FIG ). Foramen present in the sagital line between the pos- terior median chambers, just at their middle length. Postero-lateral eminences not bulging in the lachrymal region or postero-laterally, which results in a relatively shallow constriction of the interorbital region compared with the three similar species. Low sagital crest originating from back side of
eminences. Zygomatic arch thin, with dor- sal process very low and inconspicuous. Bullae relatively long and bassioccipital narrow between cochleae. Upper incisors weakly tricuspid; I 2 with well developed lingual cingulid forming a slightly excavat- ed heel; C 1 without posterior cingulum cusp.
Echolocation
Stationary bats broadcast pulses with the typical structure described for the genus Hipposideros , consisting of an initial CF segment 3–5 ms long, followed by a very short and steep linearly frequency modulat- ed sweep that falls to a frequency around
70 kHz. Almost all energy was concentrated in the second harmonic, but in some recordings a weak fundamental at half the frequency, and third and fourth harmonics were observed. CF broadcast by each bat was almost invariable, and the few varia- tions probably due to Doppler shift adjustments to the movement of the bat or sur- rounding objects. Modal CF (in kHz) for the bats recorded were: EBD 23514: 91.1; EBD 23518: 90.0; ROM 116791: 87.2; ROM 110671: 87.2; ROM 116790: 88.6; INECOL M0001: 87.5; x = 88.6. There does not appear to be sexual dimorphism in call frequencies, although the sample is small.
Comparison with most similar species
Differs from H. rotalis by: Much small- er noseleaf and internarial disc without sup- plementary leaflets. Frontal glandular sac present in females (in H. rotalis collected in the same month, males have this conspi- cuous sac, but females have it reduced to a shallow depression with some dark short hairs; however, we cannot rule out the possibility of seasonal changes). CF of the echolocation call much higher (69–71 kHz in H. rotalis ). Shorter head and bo- dy, but longer tibia and hind feet. Open- ing of the nasal channel narrower. Rostral inflations much less inflated, narrower than the toothrow (wider in H. rotalis ). Postero- lateral chamber much wider than the pos- tero-median chamber, and laterally reaching the bone bridge of the anteorbital foramen, and lacrymal chamber much reduced (the three chambers are all approximately the same width in H. rotalis — Figs. 5 View FIG and 6 View FIG ). Low sagital crest and foramen in the skull mid-line, between postero-median chambers present (both absent in H. rotalis ). Skull narrower across rostral inflations, but wider across maxillary toothrows. Ante- orbital foramen large and elongated, with bone bridge from lateral rostral inflations to zygomatic arch very thin (smaller and more rounded, with bony bridge thicker in H. rotalis ). Zygomatic arch wider and more protruding from the skull, with a more acute angle between the glenoid and the lacrymal branches.
Differs from H. orbiculus by: Smaller noseleaf and internarial disc. Frequency of the constant segment of the echolocation call higher (about 80 kHz in H. orbiculus ). Shorter head and body, and hind feet. Ears
a — Anatomical measurements for H. orbiculus taken from Francis et al. (1999), and CF data belong to Peninsular Malaya populations studied by Gould (1979)
b — H. ridleyi data are taken from Francis et al. 1999. The two CF values are the modes of the measurements for two specimens from Sabah, Borneo, Malaysia (SMF 83689–90), portrayed in Fig. 16 of that paper. Kingston et al. (2000) recorded sounds from eight specimens of H. ridleyi from Kuala Lompat, Krau Game Reserve, Pahang, Peninsular Malaysia, which had an average CF of 61.9 kHz
c — Anatomical data for H. rotalis belong to the six specimens measured by Francis et al. (1999), plus five more measured for this study field nos. AGS980508-01, from Vang Vieng area, and AGS970602-01, CMF980203-34, CMF980204-07, CMF980204-14, from Phou Khao Khouay NBCA, all from Central Lao PDR. CF data correspond to the modes of the measurements for the nine specimens portrayed in Figure 16 of Francis et al. (1999), plus the five specimens measured in this study, plus two other specimens field nos. CMF980203-34 and CMF980204-16, both from Phou Khao Khouay NBCA
d — Measurements correspond to all specimens listed in the systematic section, excepting ROM 110671 (field no. CMF980203-27)
longer. Considerably longer tail. Anterior rostral chamber of similar length to the pos- terior median rostral chambers and equal along all its width (anterior chamber longer than the posterior median chamber in the sagital edge, but narrower in the outer edge in H. orbiculus ). Postero-median chamber almost square (as an asymmetric triangle with blunt corners in H. orbiculus ). Postero- lateral chamber much wider than the pos- tero-median chamber, (the two chambers are approximately the same width in H. or- biculus). Rostral chambers with more open space and less bone density and walls (Figs. 5–6). Foramen in sagital line located in the middle of the postero-median chambers (be- hind the chambers in H. orbiculus ). Postero- lateral eminences not bulging postero-later- ally, which results in a shallower constric- tion of the interorbital region. Zygomatic arch thin and with dorsal process very low and inconspicuous as in H. rotalis (thicker and with very prominent process in H. orbi- culus and H. ridleyi ). Bullae considerably larger and bassioccipital much narrower be- tween cochleae. Canines shorter. I 2 with well developed lingual cingulid forming a slightly excavated heel (without cingulid heel in H. orbiculus ); C 1 without posterior cingulum cusp (with cingulum cusp in H. orbiculus ).
Differs from H. ridleyi by: Smaller noseleaf and internarial disc. Frequency of the constant segment of the echolocation call much higher (about 66 kHz in H. rid- leyi). Shorter hind feet. Considerably longer tail. Rostral inflations much less inflated, narrower than the toothrow (same width as toothrow in H. ridleyi ). Anterior rostral chamber of similar length to the posterior median rostral chambers and equal along all its width (anterior chamber longer than the posterior median chamber in the sagital edge, and still wider in the distal edge in H. ridleyi ). Postero-median chamber larger and almost square (almost rounded and reduced frontally in size by the expanded anterior chamber in H. ridleyi ). Only three posterior chambers (a fourth prominent chamber, larger than the median, appears between the median and the lateral chambers in H. ridleyi ). Rostral chambers with more open space and less bone density and walls. Foramen in sagital line located in the middle of the postero-median chambers (behind the chambers in H. ridleyi ). Zygomatic arch thin and with dorsal process very low and inconspicuous as in H. rotalis (thicker and with very prominent process in H. ridleyi ). Bullae considerably larger and bas- sioccipital much narrower between cochleae. Canines shorter. I 2 with well developed lingual cingulid forming a slightly excavated heel (without cingulid heel in H. ridleyi ); C 1 without posterior cingulum cusp (with cingulum cusp in H. ridleyi ). Mandible shorter. All other Hipposideros in the region have very different noseleaf mor- phology and are mostly either substantially larger or smaller.
Reproduction
The female collected in February 1998 had tiny pubic nipples and was not palpably pregnant. Of three females caught in May 1998, none was palpably pregnant, one had enlarged pubic nipples and was lactating, another had small pubic nipples, and the other had no obvious pubic nipples. Of two females collected in June 1997, none was palpably pregnant, one had enlarged pubic nipples and was lactating, while the other had no obvious pubic nipples.
Distribution
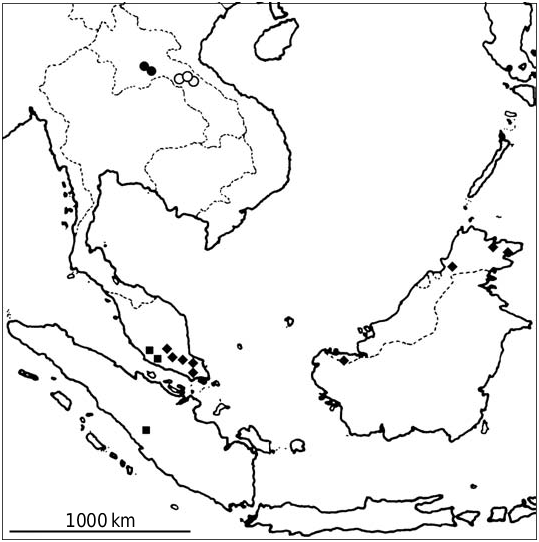
The species has been collected only in a small area at the center of Phou Khao Khouay NBCA, Saisombun Special Area, where it appeared to be relatively common, and near Ban Nampe, about 11 km NE of Vang Vieng, Vang Vieng District, Vientiane Province, all in Central Lao PDR ( Fig. 1 View FIG ). In
surveyed areas to the east the species was not observed, although H. rotalis has been caught there (Francis et al., 1999). Areas immediate to the west (Sainyabuli Prov- ince) and north (Xieng Khuang Province) of the known sites have not been surveyed. The species was not recorded in more north- ern surveyed areas (Luang Nam Tha, Luang Prabang, and Hua Phan Provinces).
Habitat
These bats have been captured in patch- es of intact and disturbed evergreen forest. In Vang Vieng there were large limestone outcrops with caves in the vicinity. Capture sites in Phou Khao Khouay were not near limestone outcrops or large caves, but the roost sites are not known. The species has been caught between 180 m (Tak Leuk waterfall, Phou Khao Khouay NBCA) and 400 m elevation (Dan Phorkeng, Phou Khao Khouay NBCA).
Morphological Comparisons and Phylo- genetic Relationships
Morphometric comparison
In general aspect and body size, H. khaokhouayensis is similar to the sympatric H. rotalis with no significant differences in forearm length, greatest skull length and mastoid width. It is also very similar to the two other species previously considered siblings of H. rotalis ( Table 1 View TABLE ). Although statistically significant for most variables measured, differences with H. rotalis are relatively small for measures related to size of wings and ears, or to length of troph- ic structures (i.e., <5% in metacarpal, tail, ear, and teeth series lengths). Instead, differ- ences in the width of trophic structures are more notable between the two species (i.e., 8–11% difference in zygomatic width, and across canines and molars widths). Of special note is the smaller size of all structures related to sound production in
H. khaokhouayensis , including width of noseleaf, internarial disk, rostral inflations, and nasal channel, and length of tympanic bulla and palate, with the exception of the bassioccipital width (the space between the bullae, which is 10% larger). The most con- spicuous differences exist in the noseleaf structure (horseshoe 23% and internarial disc 39% narrower in H. khaokhouayensis ). These differences correspond well with the large difference in the constant frequency of the echolocation calls (about 23% higher in H. khaokhouayensis ), since higher frequen- cies resonate better in shorter chords and smaller cavities ( Table 1 View TABLE ).
Although we did not perform formal tests of the morphological differences of H. khaokhouayensis with H. ridleyi and H. orbiculus , it is clear that it is very similar to them ( Table 1 View TABLE ). H. orbiculus appears more similar to H. khaokhouayensis than H. rotalis in some measurements. However there are variables and discrete characters that clearly discriminate the two species as described above, some of which point to a common ancestry of H. orbiculus and H. ridleyi (see comparison between H. kha- okhouayensis and these two species above).
Genetic analyses
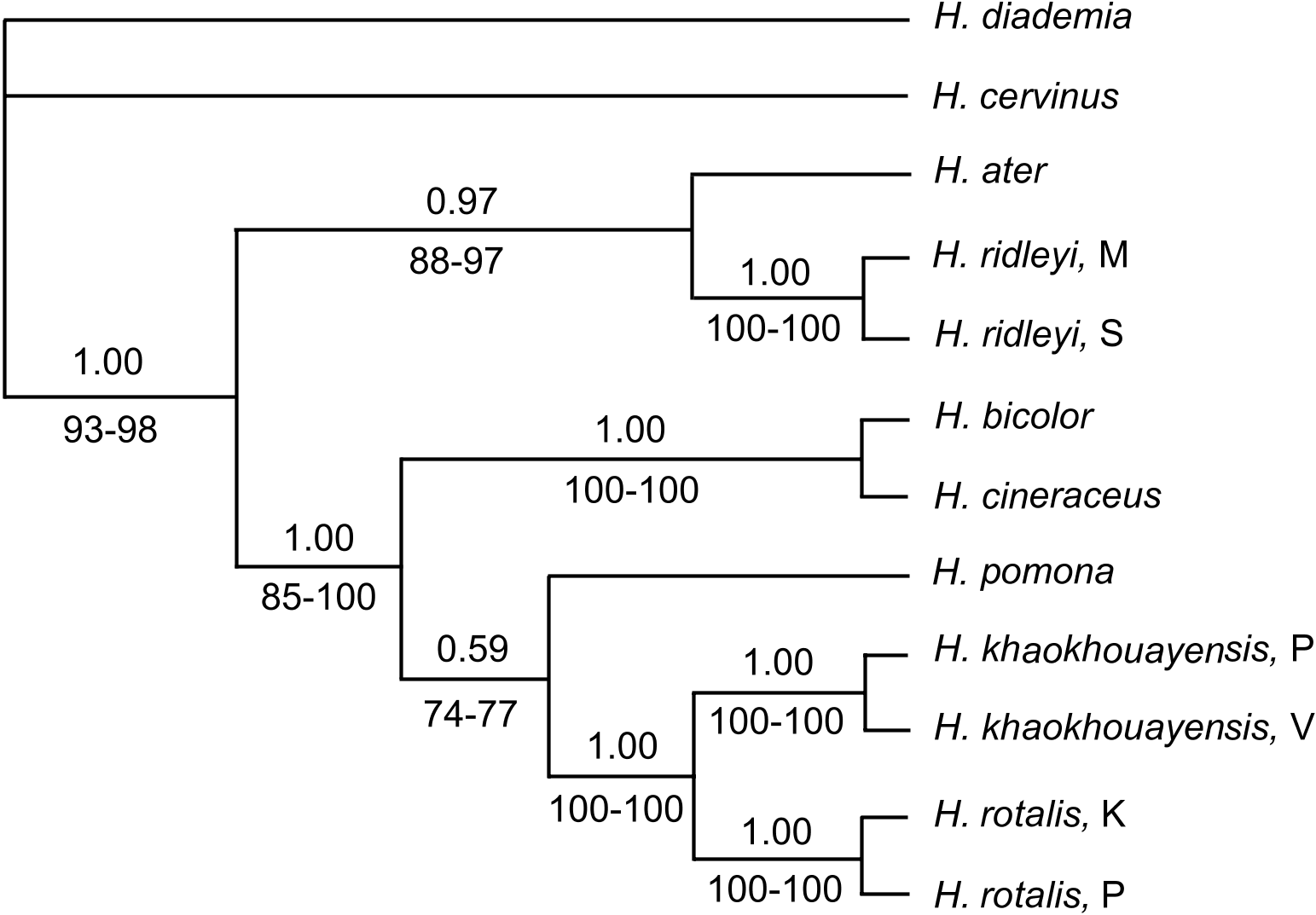
Complete nucleotide sequences of cytochrome b (1,140 bp) were generated for the 12 specimens studied (some nucleotides missing at the ends in two samples), and submitted to GenBank (Accession Numbers DQ054805 View Materials , DQ054807 View Materials – DQ054816 View Materials , and DQ219421 View Materials ). Base frequencies were similar to those reported for the gene in other species of bats with no significant differences among taxa (χ 2 = 3.66, d.f. = 33, P = 1.0), with averages of A = 0.280, C = 0.332, G = 0.137, and T = 0.251. Tamura and Nei’s (1993) equal rates genetic distances be- tween outgroup and ingroup taxa averaged 14.5% (with H. diadema ) and 14% (with
H.cervinus ). Theyranged from 7.6 to 12.7% among ingroup species, excludingH. khaokhouayensis . The shortest interspe- cific averagedistance(4.2%) wasbetweenH. rotalis and H. khaokhouayensis . Distances withinspecies ranged from 0.4% in H. khaokhouayensis to 1.7%in H. ridleyi (Table2). Of the 1,140 bp, 806(71%) sites were constant andanother126 (11%) parsimony- uninformative. Ofthe208(18% oftotalsites) informative sites 31(15%) wereat 1st-codonpositions, eight(4%) at2nd posi- tions, and 169 (81%) at 3rd positions. The datasethad astrongphylogeneticsignalsincethedistributionoftherandomtreeswasskewedsignificantlyleft (g1 = -0.90; P << 0.01). Plotsofbothtransitionsandtranversions againstTamuraandNei’s(1993) distances showedsomesaturationat thelargestdistances, butwhenallsubstitu- tions wereplottedtogether, saturationwasnotapparent. Themaximumparsimonyandtransver- sionweighted parsimonyanalysesrendered the sameunique shortest treewithlengths of576 and1,764 steps, consistencyindexes(CI) of 0.68 and 0.83, and retentionin- dexes (RI) of 0.64 and 0.81, allrespective- ly. Bootstrapfrequencieswereabove85% (97% intransversionweightedparsimony) at allnodesexceptthe onejoiningH. pomo- na assister oftheancestorofH. rotalis + H. khaokhouayensis . InallfourBayesiananalyses the chainsconvergedquickly, andreachedalikelihood asymptotebefore20,000 generations.Thetopologyof the consensustreeofthesampledtrees,afterburn-inofthepre-asymptotetrees, wasidenticaltothetreeobtainedwith the searchesunderparsimony criteria. Posteriorprobabilitieswereall above0.95, ex- ceptthenodejoiningH. pomona to thecommonancestorofH. rotalis andH. kha- okhouayensis , which appeared inonly 59% ofthetrees (Fig. 7). Members of the ingroup (all species ex- cept H. diadema and H. cervinus ) were or- ganized in two sister clades. One contained the representatives of the two geographical populations of H. ridleyi as sisters, with their ancestor sister of H. ater . The other clade contained all other specimens in the ingroup. H. bicolor and H. cineraceus ap- pear as sister species in a branch attached to the most basal node, sister to all remain- ing samples. In the crown of the clade, H. pomona appears as sister to the ancestor of H. rotalis and H. khaokhouayensis . Samples of different populations of these two species appear as reciprocally mono- phyletic. Thus, it appears that H. rotalis was not a sister species of H. ridleyi , and instead comes out as sister of H. khaokhouayensis in the best phylogenetic hypothesis obtained under the different optimisation criteria used ( Fig. 7 View FIG ). The hypothesis of monophyly of H. ridleyi with H. rotalis and H. kha- okhouayensis was rejected since no topolo- gy with a taxa partition including these three species only was found in the 99.9% credible set of trees.



| ROM |
Canada Entomology Department, Royal Ontario Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.