
Vosmaeropsis recruta, Cavalcanti, Fernanda F., Bastos, Nilma & Lanna, Emilio, 2015
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3956.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:20A373F2-27BE-4599-A586-C78F7378ED9F |
|
DOI |
https://doi.org/10.5281/zenodo.5685379 |
|
persistent identifier |
https://treatment.plazi.org/id/03901754-F50F-FFBC-FF6A-883F7698F953 |
|
treatment provided by |
Plazi |
|
scientific name |
Vosmaeropsis recruta |
| status |
sp. nov. |
Vosmaeropsis recruta View in CoL sp. nov.
Etymology. From Portuguese recruta = recruit (noun). The name was selected because the specimens were sampled on artificial substrates used to the recruitment of marine invertebrate larvae.
Diagnosis. Vosmaeropsis with leuconoid type of aquiferous system. The shape is tubular, and the sponge body is very hispid due to several large diactines protruding through the cortex. In these diactines the distal tip is commonly arrow-shaped. The choanoskeleton is formed by two categories of triactines, one of them is subatrial. Atrial skeleton is composed mainly of triactines and few tetractines, occasionally with tangential large diactines.
Type material. UFBA 4456-POR (Holotype), UFBA 4457-POR, and UFBA 4458-POR (Paratypes). Forno Harbour, Arraial do Cabo, Rio de Janeiro, Brazil; depth: 1 m; 17/VII/2009; collected by F.F. Cavalcanti.
Type locality. Arraial do Cabo, Rio de Janeiro, Brazil.
Additional material. UFBA 4469-POR, UFBA 4470-POR, and UFBA 4471-POR. From the same locality of the type specimens.
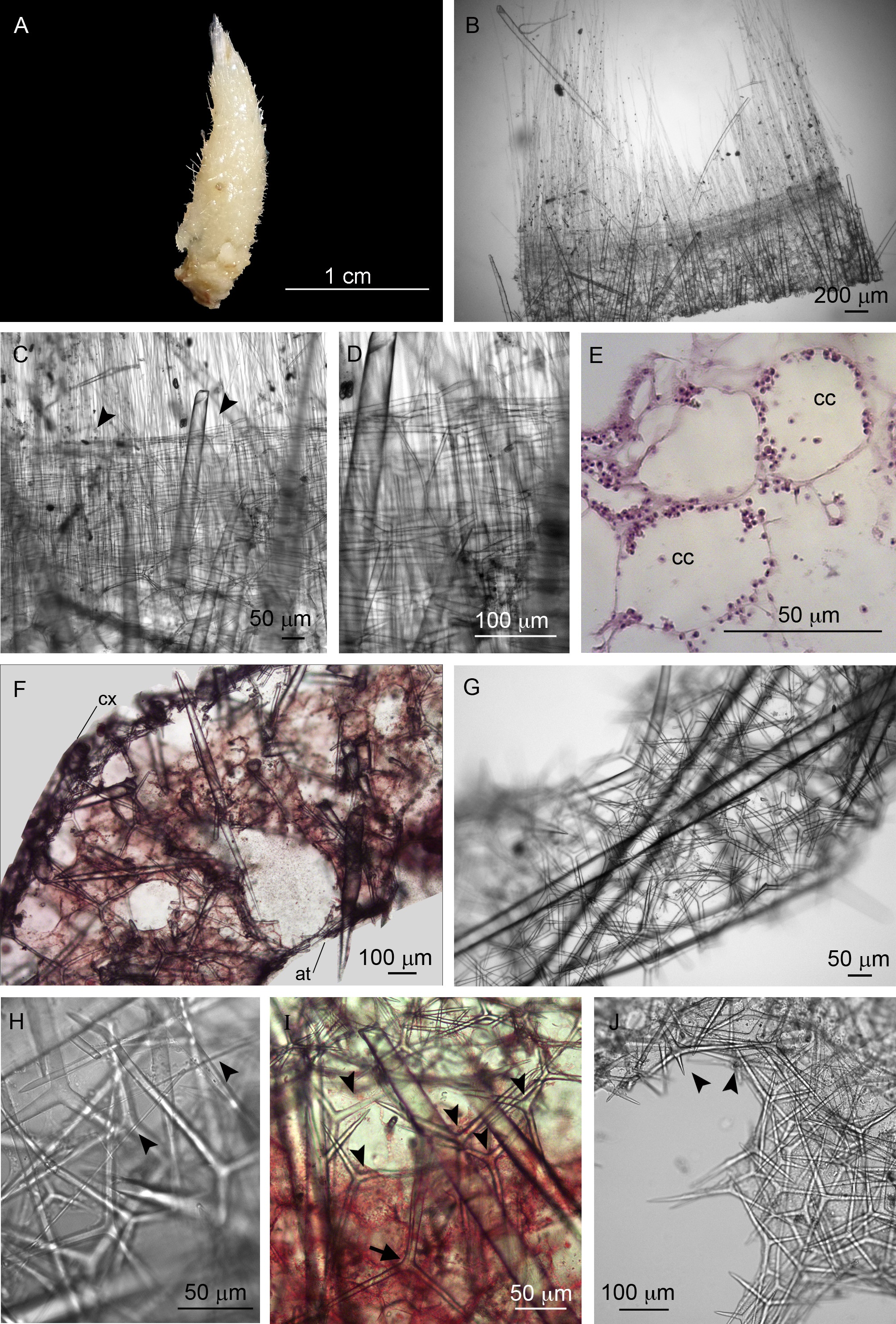
Description. Colour is beige, alive and after fixation ( Figure 2 View FIGURE 2 A). The holotype is slightly larger than the paratypes, and it measures 2.0 x 0.8 cm (height x width). The sponge is tubular, with an apical osculum ornamented by a fringe of trichoxea ( Figures 2 View FIGURE 2 A–B). Sagittal triactines surround the base of the fringe, forming a ring ( Figures 2 View FIGURE 2 B–D). The surface is very hispid due to the presence of several large diactines with the distal tips protruding through the cortex. These spicules are mainly perpendicular to the sponge body, except near the osculum where they are tangential. The atrial cavity is wide. The aquiferous system is leuconoid, with spherical choanocyte chambers ( Figure 2 View FIGURE 2 E).
Large diactines cross the sponge wall and sometimes reach the atrial cavity ( Figure 2 View FIGURE 2 F). The cortical skeleton is composed of tangential triactines ( Figure 2 View FIGURE 2 G). Fragments of trichoxea were also found in this region. They are scarce and tangential to the cortex, but without organisation ( Figure 2 View FIGURE 2 H). These spicules also occur at the atrial skeleton. Below the cortex, the subcortical region is formed by a layer of pseudosagittal triactines, which point one of their paired actines toward the atrium ( Figure 2 View FIGURE 2 I). Large triactines are present at the choanosome. These spicules are dispersed without organisation, but some of them point the unpaired actine towards the cortex, forming an inarticulated skeleton ( Figure 2 View FIGURE 2 I). Rare tetractines were found surrounding an exhalant canal. Subatrial triactines were observed, but they are not abundant. The atrial skeleton is formed mainly by triactines and few tetractines ( Figure 2 View FIGURE 2 F). Rare tangential diactines similar to those found on the cortex ( Figure 2 View FIGURE 2 G) are also present at the atrial skeleton of the paratypes (UFBA 4457-POR and UFBA 4458-POR).
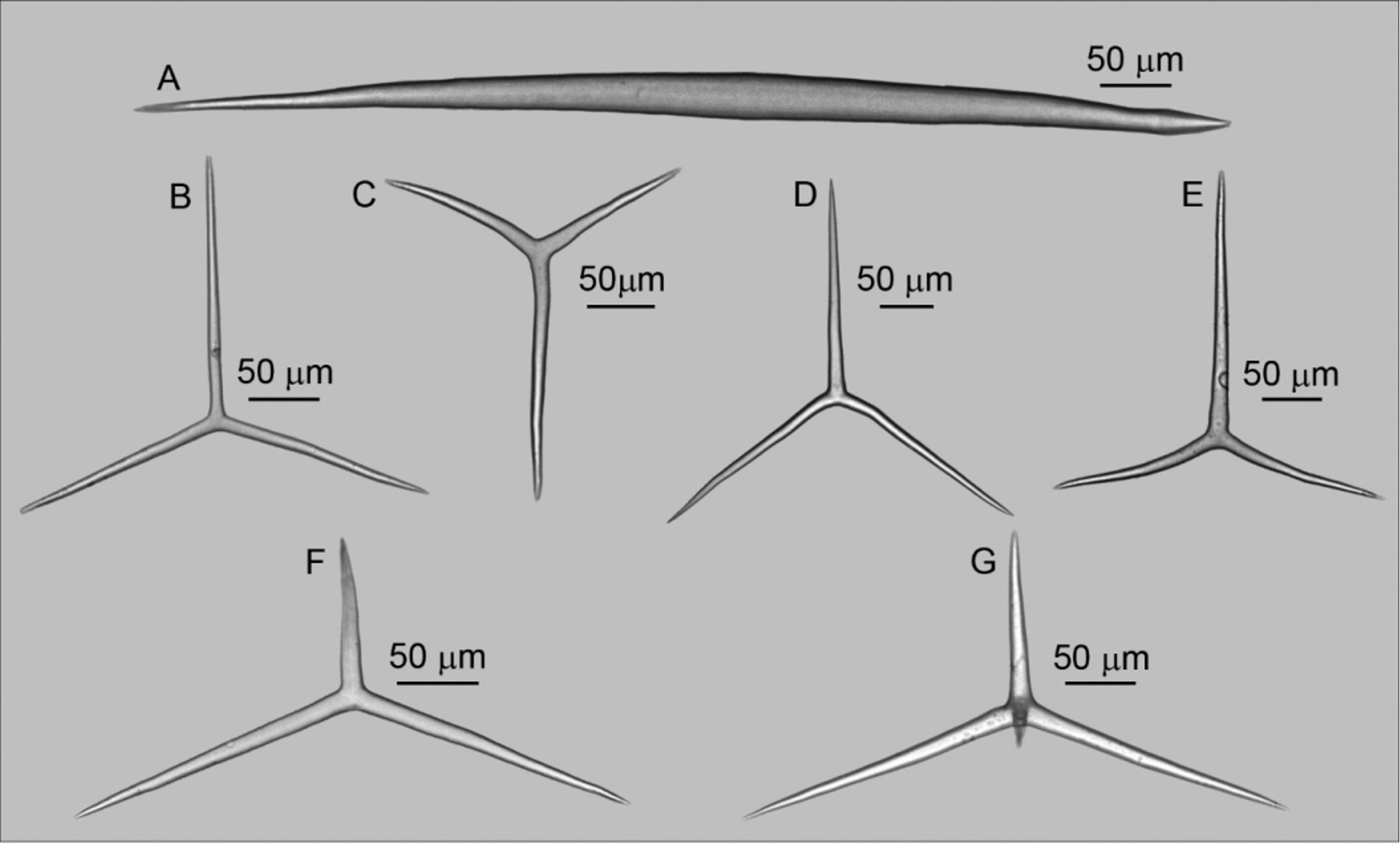
Spicules ( Table 1): Trichoxeas ( Figure 2 View FIGURE 2 H): Very thin spicules. They are usually broken, but measure at least 150 µm.
Diactines ( Figure 3 View FIGURE 3 A): Large and stout. The proximal tip is sharply pointed but the distal one can be sharp or arrow-shaped [372–949.1 ± 59.7–2156/ 20–40.6 ± 2.7–78 µm (n= 3 specimens)];
Cortical triactines ( Figure 3 View FIGURE 3 B): Almost regular and variable in size. Actines are straight, cylindrical, and with blunt tips. The unpaired actine can be slightly larger or slightly shorter than the paired ones [Paired actines: 110– 186.7 ± 21.3–300/ 10–11.3 ± 0.5–20 µm; unpaired actine: 90–181.9 ± 30.7–280/ 10–11.6 ± 0.5–20 µm (n= 3 specimens)];
Subcortical triactines ( Figure 3 View FIGURE 3 C): Pseudosagittal. Actines are slightly conical and blunt. The longest paired actine is commonly straight while the shortest one is curved. The unpaired actine is commonly short and straight [Paired actine I: 80–169.4 ± 28.3–300/ 10–14.3 ± 3.8–20 µm; paired actine II: 150–231.0 ± 23.7–330/ 10–14.5 ± 0.8–25 µm; unpaired actine: 70–145.2 ± 23.2–270/ 10–14.6 ± 0.4–20 µm (n= 3 specimens)];
Choanosomal triactines ( Figure 3 View FIGURE 3 D): Actines are slightly conical and equiradiated, with blunt tips. The paired actines (both or only one of them) are curved to the opposite side of the unpaired one. Additionally, one of the paired actines is commonly wavy. They are larger than the other triactine categories [Paired actine: 140–236.4 ± 29.7–340/ 10–15.4 ± 0.5–20 µm; unpaired actine: 120–224.8 ± 35.4–340/ 10–16.0 ± 1.4–20 µm (n= 3 specimens)]; Subatrial triactine ( Figure 3 View FIGURE 3 E): Sagittal. Actines are slightly conical and with blunt tips. The paired actines are curved and are commonly shorter than the unpaired one [Paired actine: 130–203.6 ± 12.6–280/ 15–16.3 ± 0.2–20 µm; unpaired actine: 190–245.6 ± 33.3–350/ 15– 17.7 ± 0.2–20 µm (n= 3 specimens)].
Atrial triactines ( Figure 3 View FIGURE 3 F): They are sagittal. Actines are slightly conical, straight, with blunt tips. The unpaired actine is shorter than the paired ones [Paired actines: 120–216.6 ± 29.0–310/ 10–13.0 ± 0.8–20 µm; unpaired actine: 50–117.1 ± 9.1–190/ 10– 13.0 ± 0.5–20 µm (n= 3 specimens)];
Atrial tetractines ( Figures 3 View FIGURE 3 G; 4): They are similar to the atrial triactines. The apical actine is short, smooth and straight [Paired actines: 100–220.7 ± 33.7–320/ 10–18.5 ± 1.1–20 µm; unpaired actine: 50–132.8 ± 4.7–220/ 10– 15.1 ± 0.5–20 µm; apical actine: 20–45.4 ± 10.2–80/ 10–10.6 ± 1.7–20µm (n= 3 specimens)].
Spicule Actine Length (µm) Width (µm) N
Min. Mean SD Max. Mean SD
Diactines 392 905.9 418.0 2156 37.9 7.4 23 Cortical triactines Paired 110 162.7 25.2 220 10.8 1.9 30 Unpaired 130 165.3 23.3 220 11.0 2.0 30 Subcortical triactines Paired 1 80 141.3 30.4 200 12.7 2.5 30 Paired 2 160 209.3 20.5 250 12.8 2.8 30 Unpaired 100 126.0 19.8 170 13.0 2.5 30 Choanosomal triactines Paired 160 208.0 25.8 250 15.0 1.9 30 Unpaired 140 199.7 40.2 280 15.0 0.0 30 Subatrial triactines Paired 130 189.2 23.5 220 16.4 2.2 12 Unpaired 200 222.9 20.9 260 18.4 2.1 12 Atrial triactines Paired 120 201.0 31.4 250 12.5 2.9 30 Unpaired 60 107.7 27.4 180 12.3 2.9 30 Atrial tetractines Paired 150 197.0 29.0 270 14.7 1.8 30 Unpaired 80 131.9 37.2 210 14.8 1.7 26 Apical 40 49.0 6.4 60 10.2 1.1 20 Ecology. All the specimens were found on plastic structures used as artificial substrate to the settlement of larvae of marine invertebrates. As these structures remained submerged for two months, this is the maximum age of the specimens.
Remarks. Vosmaeropsis is currently formed by 21 species. A remarkable character of V. recruta sp. nov. is the presence of large diactines in its skeleton, which also occur in ten other species of the genus ( Table 3). These species are: V. cyathus ( Verril, 1873) , V. g r i s e a Tanita, 1939, V. h i s p an i ca Ferrer-Hernández, 1933, V. inflata Tanita, 1942 , V. japonica Hôzawa, 1929 , V. macera ( Carter, 1886) , V. mackinnoni Dendy & Frederick, 1924 , V. oruetai Ferrer-Hernández, 1918 , V. sericata and V. spinosa Tanita, 1943 . Vosmaeropsis recruta sp. nov. can be easily differentiated from most of these species by its skeletal composition. For example, its atrial skeleton is formed by tri- and tetractines, while V. macera and V. mackinnoni have only atrial triactines, and V. hispanica , V. or u e t a i, V. sericata and V. s pi n os a have only atrial tetractines. The atrial skeleton can also be used to differentiate V. recruta sp. nov. from V. japonica . In the latter, it is composed mainly by the paired actines of the subatrial spicules, which does not occur in V. recruta sp. nov. The main difference between V. recruta sp. nov. and V. cyathus is the absence of choanosomal tetractines in the latter and the abundance of diactines in V. recruta sp. nov. (diactines are few in V. cyathus ). Finally, in relation to V. inflata , the new species can be easily differentiated by its surface type and arrangement of the diactines—in V. inflata these spicules never protrude from the surface while in V. recruta sp. nov. they do, making it strongly hispid. Thus, among the Vosmaeropsis species that have large diactines, the most similar species to V. recruta sp. nov. is V. g r i s ea, which occurs on the Japanese coast ( Tanita, 1939). The main differences between them are the shape of the diactines (pointed at both sides in V. grisea and arrow-shaped in V. recruta sp. nov.) and the shape and size of the atrial tri- and tetractines (the unpaired actine is longer than the paired actines in V. grisea , while in V. recruta sp. nov. the opposite occurs). The comparison between V. recruta sp. nov. and V. complanatispinifera sp. nov. is provided below.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.