
Opacuincola mete kahurangi, Verhaegen & Haase, 2021
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.731.1205 |
|
publication LSID |
lsid:zoobank.org:pub:CBF3D6E0-9896-4852-AA9F-6DD49C950795 |
|
DOI |
https://doi.org/10.5281/zenodo.4446922 |
|
persistent identifier |
https://treatment.plazi.org/id/D05FF1E3-5A77-4C4F-AA94-C8E7356F36C7 |
|
taxon LSID |
lsid:zoobank.org:act:D05FF1E3-5A77-4C4F-AA94-C8E7356F36C7 |
|
treatment provided by |
Plazi |
|
scientific name |
Opacuincola mete kahurangi |
| status |
subsp. nov. |
Opacuincola mete kahurangi subsp. nov.
urn:lsid:zoobank.org:act:D05FF1E3-5A77-4C4F-AA94-C8E7356F36C7
Figs 4 View Fig G–H, 5 View Fig F–G, 6 View Fig E, 11 View Fig C, 13 View Fig ; Table 1 View Table 1
Diagnosis
The new subspecies differs from the nominate form in shell morphology. It is much larger and broadlyconical while the latter has a rather ovate shell. Both are identical in anatomy.
Etymology
Opacuincola mete kahurangi subsp. nov. is named after Kahurangi National Park, the second largest National Park in New Zealand in the northeast of the South Island, where the type locality is situated.
Material examined
Holotype ( Fig. 4G View Fig ) NEW ZEALAND • Kahurangi National Park , NE of Karamea, Fenian Track, under bridge over Cavern Creek; 41°12ʹ02.5ʺ S, 172°11ʹ08.4ʺ E; 5 Mar. 2016; G. Verhaegen and M. Haase leg.; on rough rocks in river; NMNZ.M.330193 . GoogleMaps
Paratypes ( Figs 4H View Fig , 5 View Fig F–G) NEW ZEALAND • 21 specs; same collection data as for holotype; NMNZ.M.330194 GoogleMaps .
Description
SHELL ( Figs 4 View Fig G–H, 5F–G). Broadly-conical to globular, only about 1.2 times as high as than wide, translucent with brown periostracum; protoconch almost smooth, comprising ca 1 whorl ( Fig. 6E View Fig ); entire shell with 3.125 to 4 whorls, teleoconch without structure apart from growth lines; umbilicus a wider slit; aperture large, wider than high, about half as high as total shell height and more than half as wide as total width, apertural lip slightly sinuated both ad- and abapically.
OPERCULUM. Orange, paucispiral; nucleus submarginal, without peg.
EXTERNAL FEATURES. Epidermis without pigment; eyes well developed and entirely pigmented; tentacles without particular ciliation.
MANTLE CAVITY (n = 4). 9–12 ctenidial filaments; osphradium ovate-elongate, behind middle of gill.
DIGESTIVE SYSTEM. Radula has formula R 5-6 1 5-6/3-4 3-4, L 5 1 5-6, M1 23-28, M2 29-30; stomach without caecum; rectal loop pointing left in roof of mantle cavity, in males more distinct than in females.
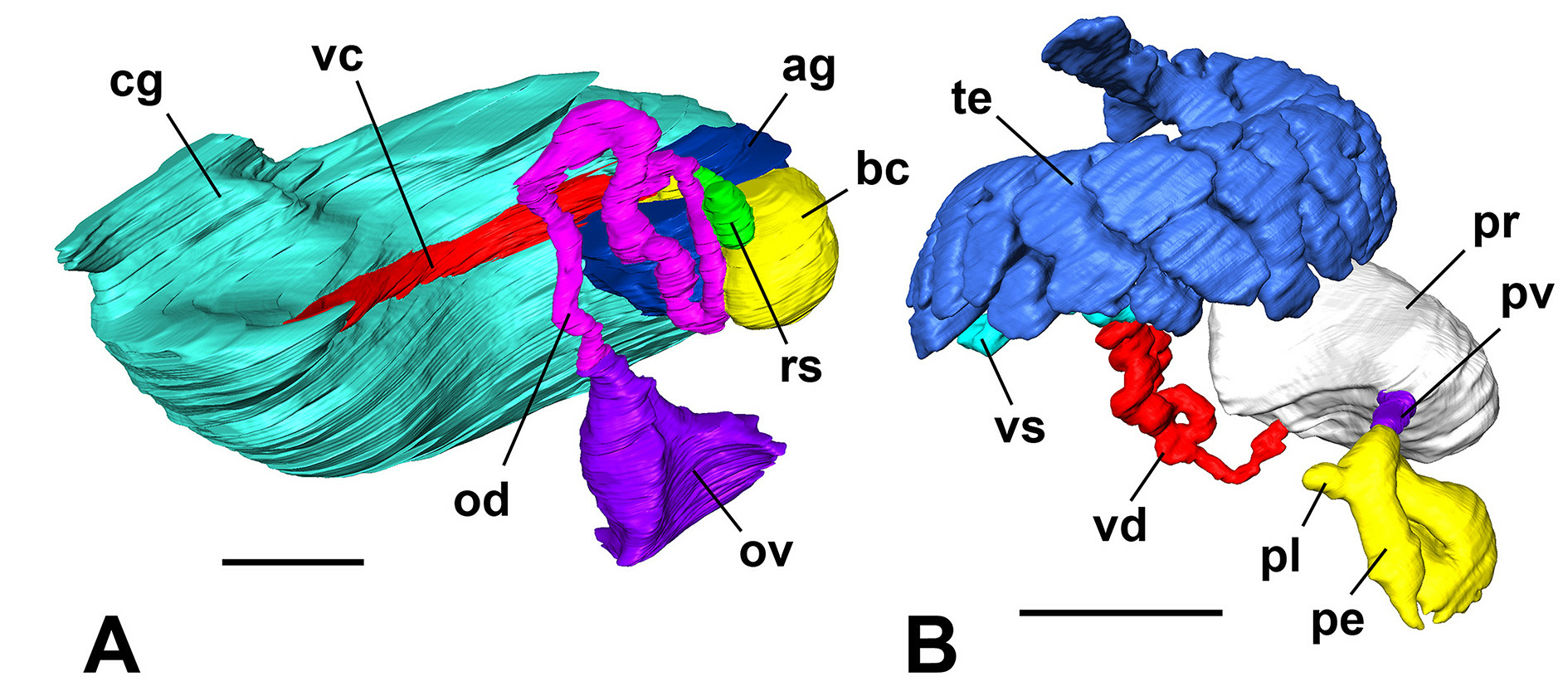
FEMALE GENITALIA (n = 3; Fig. 13A View Fig ). Ovary small, simple sac starting ca 1 whorl below apex, comprising 0.25 whorl and not reaching stomach; renal oviduct first coiling 180° clockwise, then 270° counterclockwise; one distal, large receptaculum seminis lying against anterior area of bursa copulatrix; bursa copulatrix large, globular, largely behind short albumen gland, fairly long bursal duct entering anteriorly; ovoviviparous, brooding at least four embryos in pallial oviduct, pallial oviduct as brood pouch with very short albumen gland and large capsule gland, the latter histologically uniform in CT scans.
MALE GENITALIA (n = 3; Fig. 13B View Fig ). Testis lobate sac, starting ca 1 whorl below apex, comprising up to 0.75 whorl, not reaching stomach; vesicula seminalis coils along anterior third of testis; proximal vas deferens inserts close to middle of kidney-shaped prostate, distal vas deferens leaving anteriorly; penis not very massive, with subterminal swelling and short terminal filament, fairly long muscular lobe on right side eventually bent forward ( Figs 11C View Fig , 13B View Fig ).
Remarks
In shell morphology, Op. m. kahurangi ssp. nov. is clearly distinct form the nominate form. Univariate tests could not be conducted because of the small sample size available for the latter. However, the PCA is unambiguous ( Fig. 3 View Fig ). Anatomically, they are practically identical and in the mitochondrial phylogeny not separable ( Fig. 2 View Fig ). Both are known each from a single locality, which are ca 90 km apart (as the crow flies). Because of the high overall similarity, we distinguish these forms only as subspecies, although they can easily be told apart by their shells. However, we do not know if there exist connecting populations between both sites mediating gene flow in a stepping stone-like fashion. Considering similar cases among Tateidae form New Zealand and other places, we might be witnessing species in statu nascendi or already be dealing with genetically incompatible but mitochondrially undifferentiated, young species (e.g., Haase 2005, 2008; Zielske & Haase 2014a, 2014b). Nuclear genetic data might be more informative in this ambiguous situation. The new subspecies represents one of the rare cases where a small tateid is not restricted to springs or small streams. Due to the unresolved situation in the mitochondrial phylogeny ( Fig. 2 View Fig ), it is not reasonable to list diagnostic alignment positions.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |