
Lophoplax sculpta ( Stimpson, 1858 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5244.5.2 |
|
publication LSID |
lsid:zoobank.org:pub:B55396DA-15DB-4CDF-81FD-78686A0EC3A2 |
|
DOI |
https://doi.org/10.5281/zenodo.7663756 |
|
persistent identifier |
https://treatment.plazi.org/id/039087CD-7E07-7320-47C6-FAA7F6E7FCE3 |
|
treatment provided by |
Plazi |
|
scientific name |
Lophoplax sculpta ( Stimpson, 1858 ) |
| status |
|
Lophoplax sculpta ( Stimpson, 1858) View in CoL
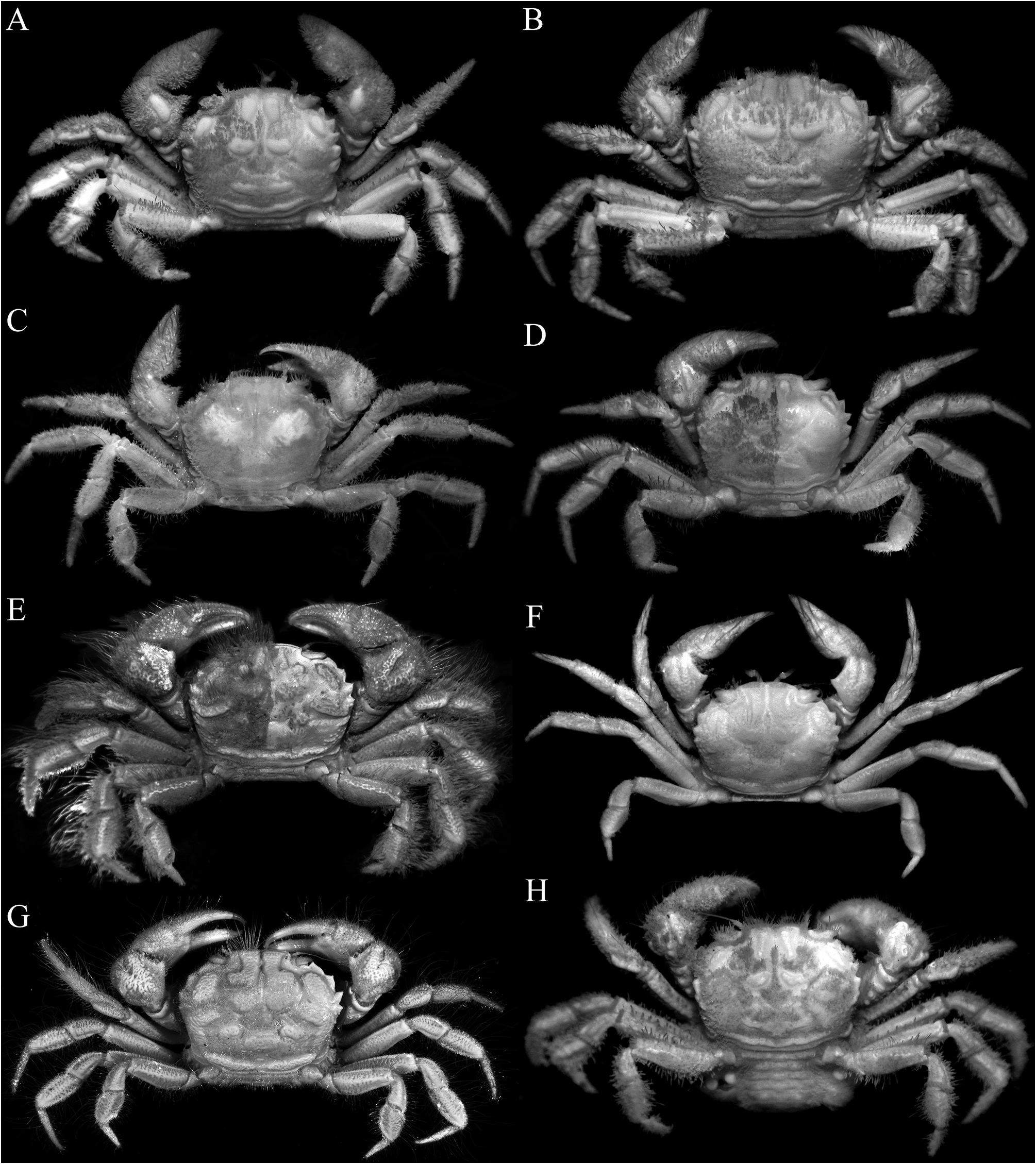
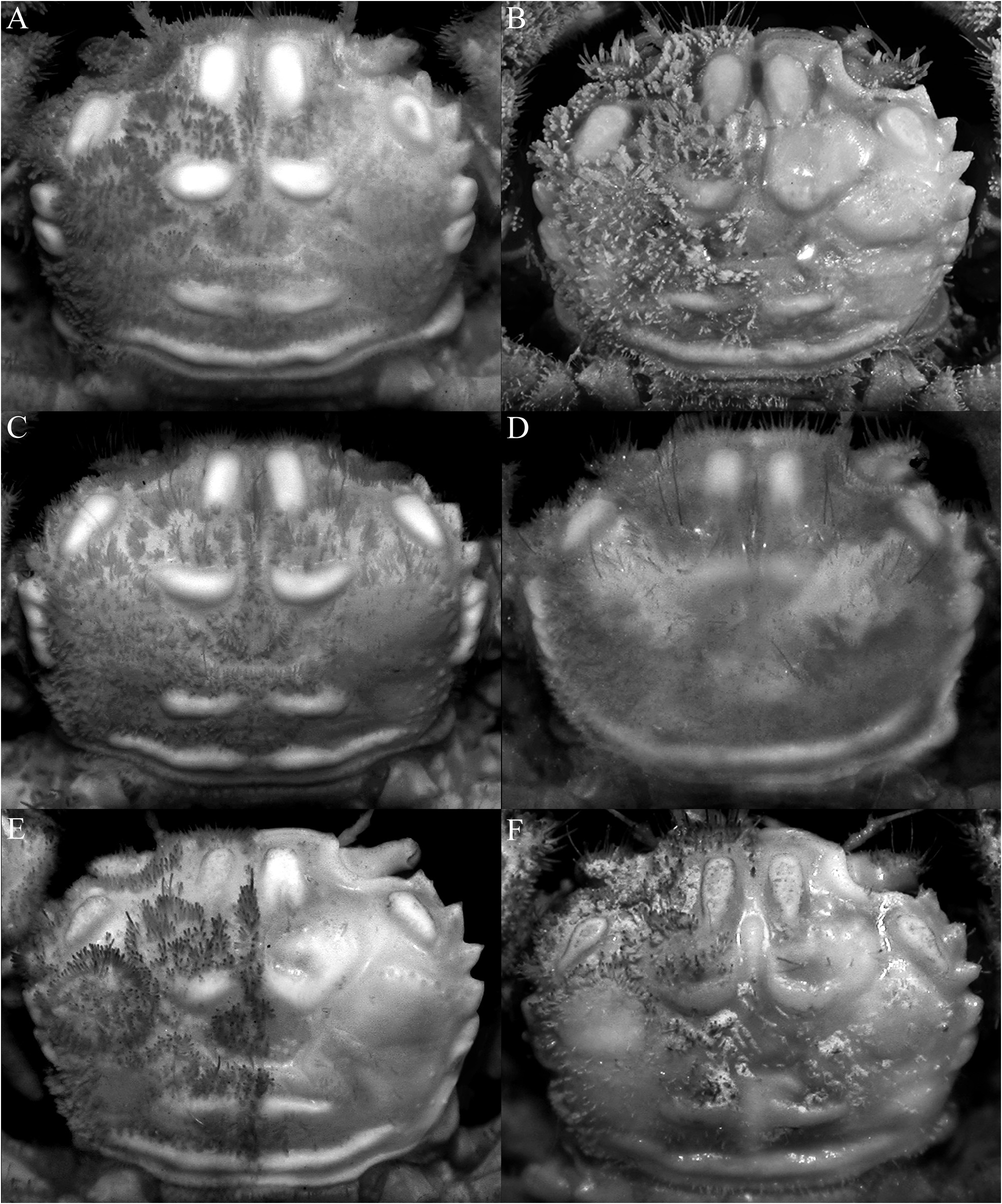
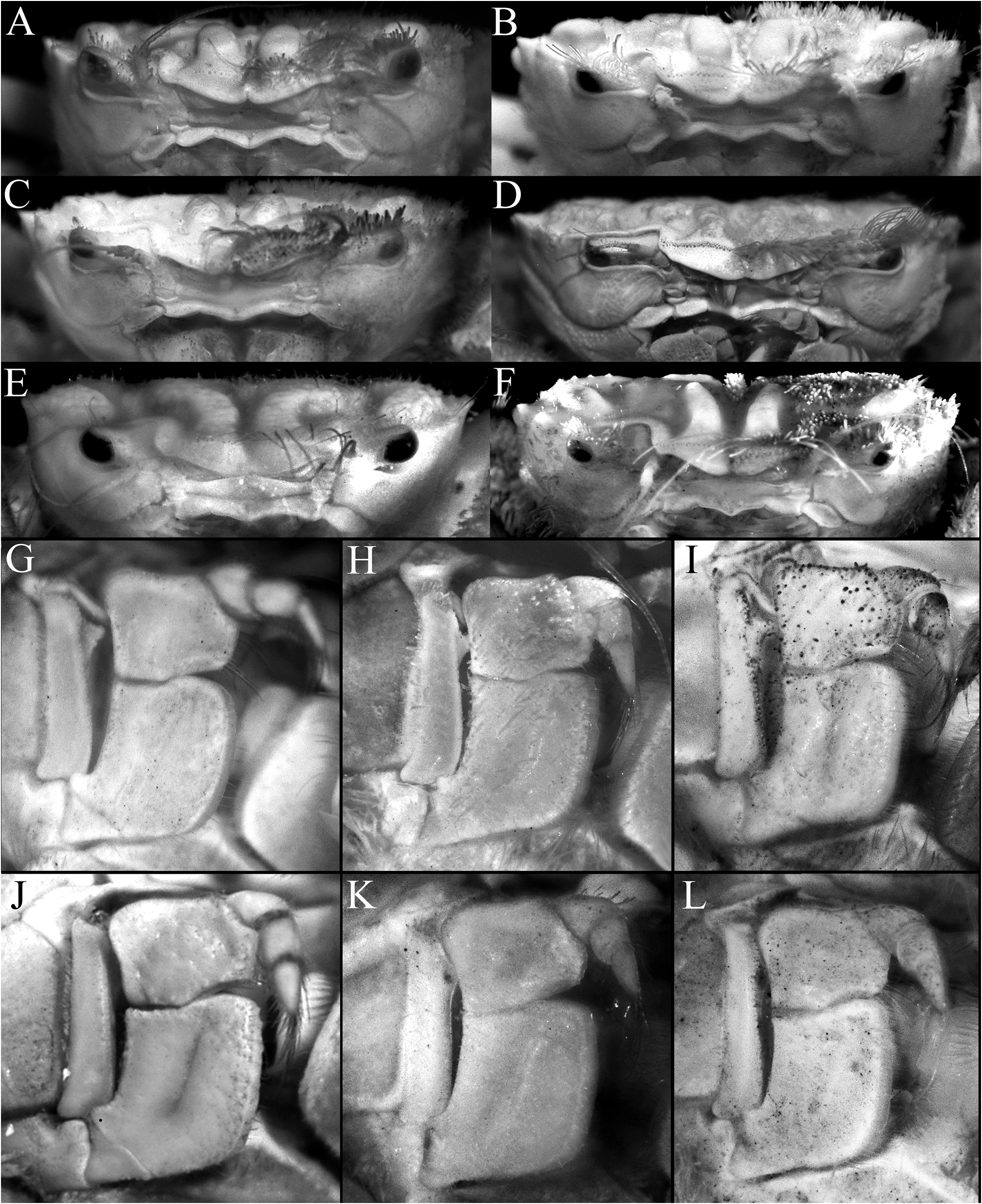
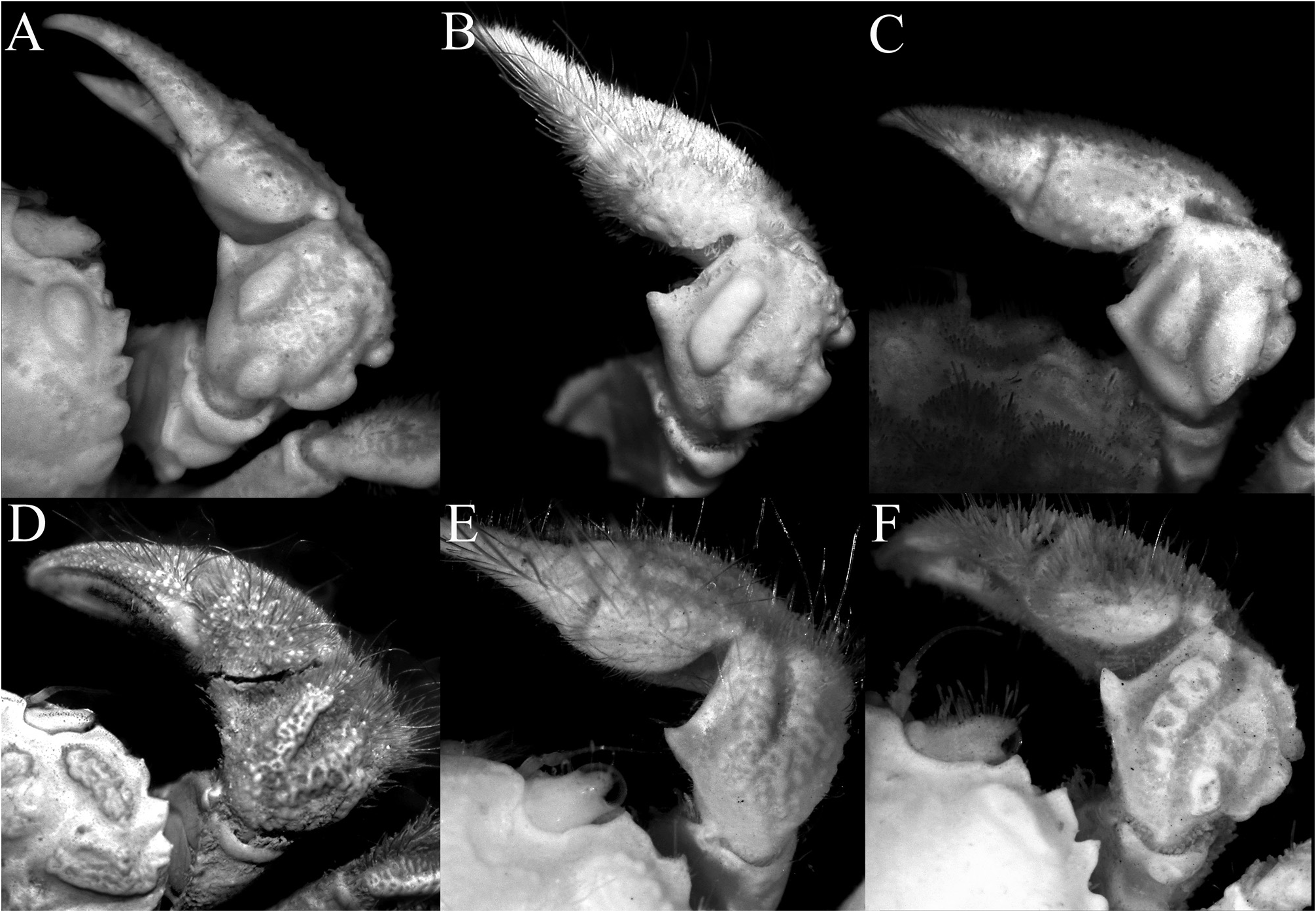
( Figs. 1A, B View FIGURE 1 , 2A, B View FIGURE 2 , 3A–C View FIGURE 3 , 5A, B, G, H View FIGURE 5 , 6A, B View FIGURE 6 , 7A, B, G, H View FIGURE 7 , 8A, B View FIGURE 8 , 9A–D View FIGURE 9 , 10A–E View FIGURE 10 )
Pilumnoplax sculpta Stimpson, 1858: 93 ; Miers 1886: 226; Stimpson 1907: 91, pl. 11 fig. 3.
Lophoplax sculpta View in CoL — Tesch 1918: 199; Sakai 1939: 567; Serène 1968: 91; Serène & Lohavanijaya 1973: 69, 75; Serène et al. 1974: 24; Serène et al. 1976: 19; Sakai 1976: 304, 541; Takeda 1977: 120, fig. 1; Miyake 1983: 231; Dai et al. 1986: 379, fig. 200 (1, 2), pl. 55 (4); Ng 1987: 79, 100, 101; Nagai & Nomura 1988: 196; Dai & Yang 1991: 409, fig. 200 (1, 2), pl. 55 (4); Nomura et al. 1996: 17; Sakai 2003: 22; Marumura & Kosaka 2003: 47; Ng et al. 2008: 143, fig. 111; Castro & Ng 2010: 25; Naruse 2010: 31; Maenosono 2019: 30, figs. 8A, 9, 13A, 14I, J.
Type material. Neotype (here designated): male (7.8 × 6.3 mm) (RUMF-ZC-8127), Minatagawa, Urasoe City, Okinawa, Japan, coll. T. Maenosono, 6 January 2008.
Other material examined. Japan — 1 male (7.8 × 6.2 mm) ( ZRC 2022.777 View Materials ), Ukaji, Onna Village, Okinawa Island, Ryukyu Islands , coll. T. Maenosono, 22 August 2009 . Philippines — 1 female (10.4 × 8.5 mm) ( ZRC 2019.1611 View Materials ), station M18, sandy bottom with seagrass, Gak-Ang Island , Panglao, Bohol, 0–1 m, coll. PANGLAO 2004 Expedition, 10–12 June 2004 .
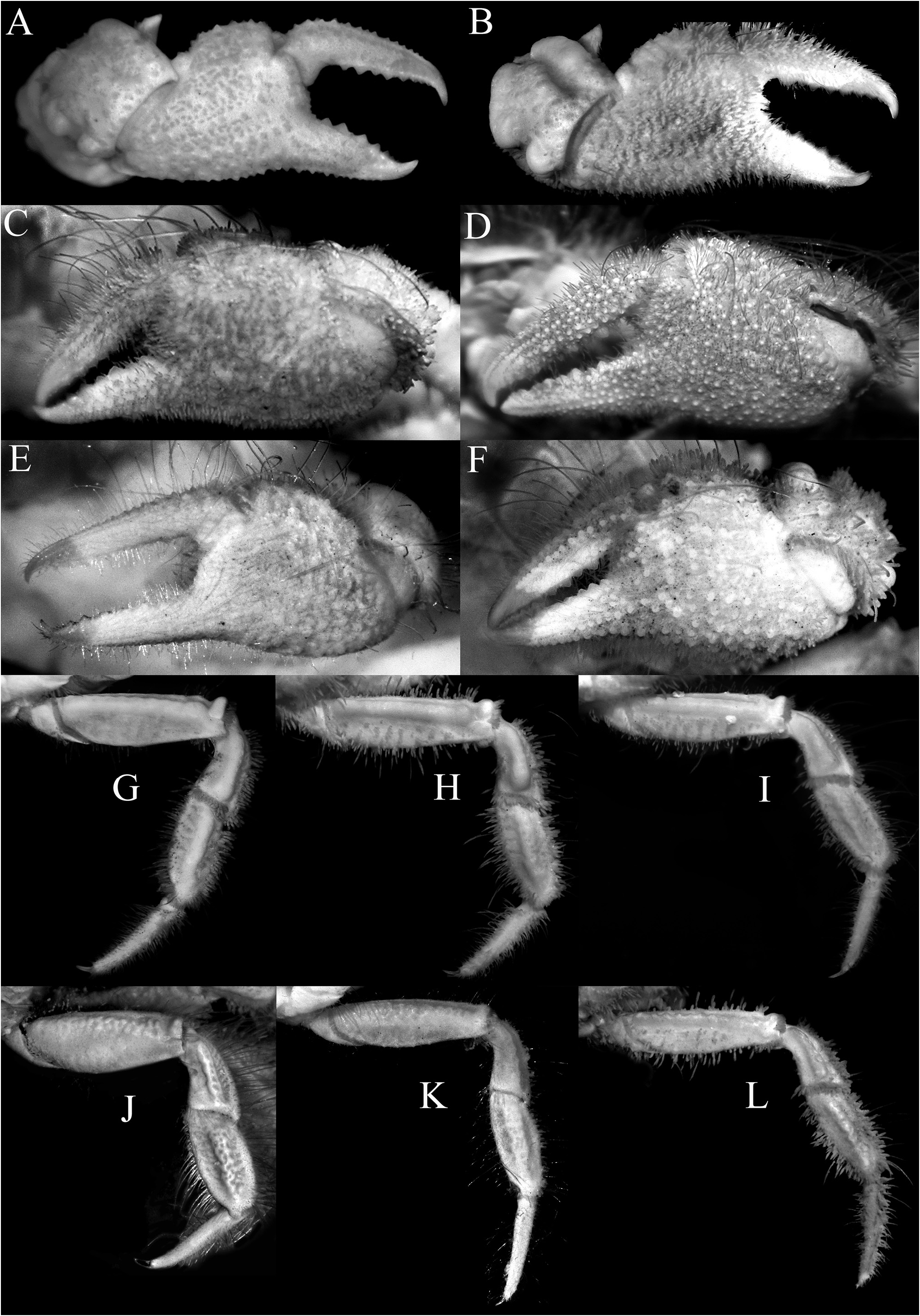
Diagnosis. Carapace subquadrate ( Fig. 3A–C View FIGURE 3 ); epigastric, protogastric, hepatic, cardiac and intestinal areolets all glabrous, prominent, smooth; epigastric areolets relatively wide, ovate, not fused with protogastric areolets ( Fig. 3A–C View FIGURE 3 ); protogastric areolet prominent, transversely ovate to weakly crescent-shaped, positioned transversely ( Fig. 3A–C View FIGURE 3 ); cardiac areolet transverse in position ( Fig. 3A–C View FIGURE 3 ); hepatic areolet swollen, prominent, sometimes with a median depression, proportionately short, reaching only to base of first anterolateral tooth ( Fig. 3A–C View FIGURE 3 ); external orbital tooth acutely triangular ( Fig. 3A–C View FIGURE 3 ); anterolateral teeth sharply pointed ( Fig. 3A–C View FIGURE 3 ); last anterolateral tooth rounded, at junction between antero- and posterolateral margins, partially fused with 1 or 2 partially fused rounded to subtruncate tubercles each often with median depression ( Figs. 3A–C View FIGURE 3 , 9A View FIGURE 9 ); epistome relatively wide longitudinally ( Fig. 5A, B View FIGURE 5 ); median lobe of posterior margin of the epistome broadly triangular, with lateral margins deeply concave ( Fig. 5A, B View FIGURE 5 ). Third maxilliped with merus quadrate, ischium subrectangular ( Fig. 5G, H View FIGURE 5 ); dorsal and lateral surfaces of carpus of cheliped with 5 distinct smooth tubercles ( Fig. 6A, B View FIGURE 6 ). Cheliped fingers not distinctly bent, pollex subparallel with the ventral margin of palm ( Fig. 7A, B View FIGURE 7 ). P2–P5 not prominently elongate ( Figs. 2A, B View FIGURE 2 , 7G, H View FIGURE 7 ); outer surface of the P3–P5 merus, carpus and propodus relatively smooth to slightly rugose ( Fig. 6A, B View FIGURE 6 ). Male pleonal somites 4–6 transversely narrow, telson slightly longer than broad, triangular ( Figs. 9B, D View FIGURE 9 , 10A View FIGURE 10 ). G1 strongly sinuous, distal part long, gently curved, elongate ( Fig. 10B–D View FIGURE 10 ).
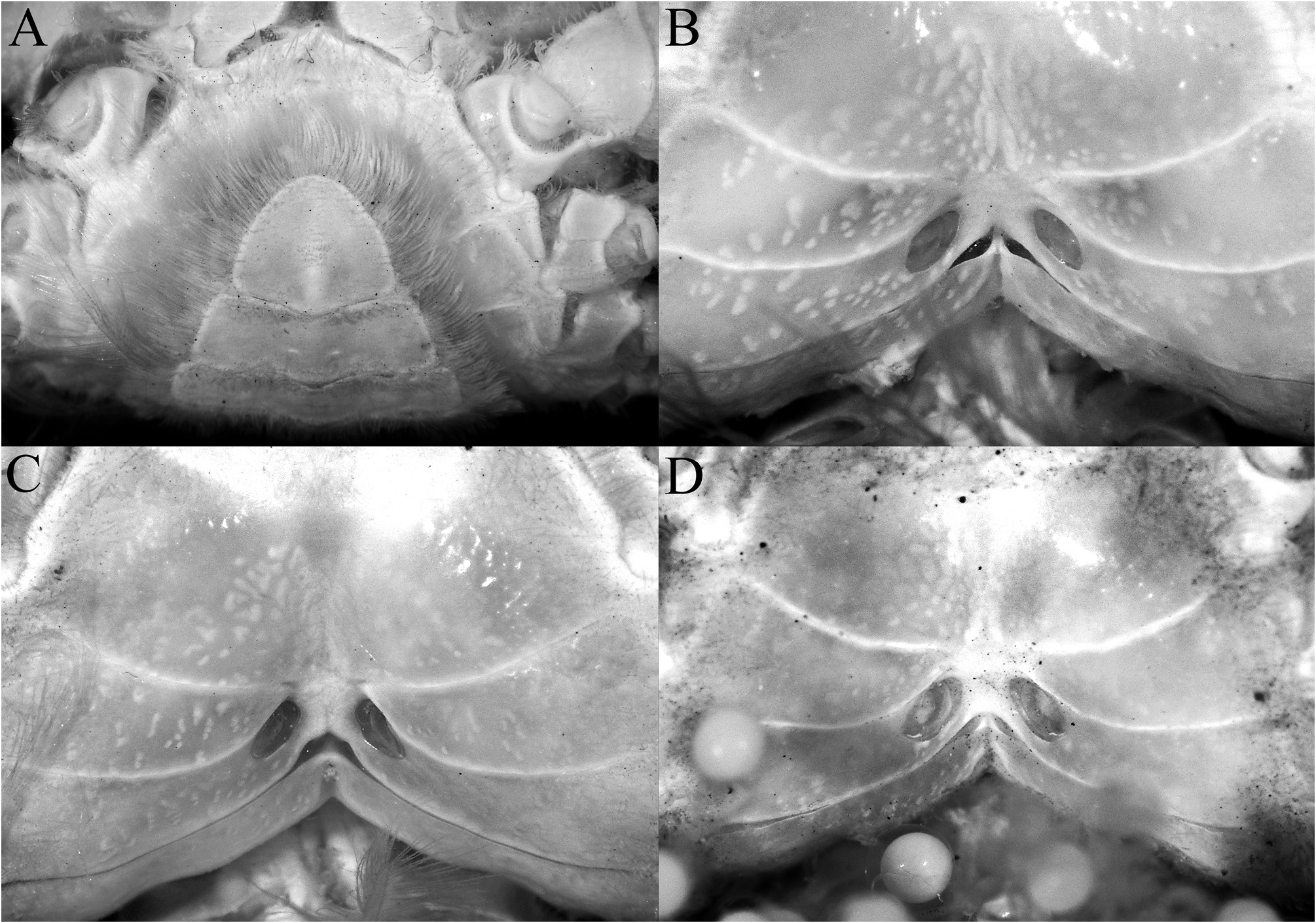
Description of male. Carapace subquadrate, slightly broader than long; dorsal surface gently convex in anterior half, relatively flatter on posterior half; regions distinct; surface entirely covered by short, thick setae and scattered longer plumose setae, forming spongy tomentum except for smooth, glabrous areolets on symmetrical epigastric, protogastric, hepatic, cardiac and intestinal regions; epigastric region relatively wide, longitudinally subrectangular, not fused with protogastric areolets; protogastric region transversely ovate to weakly crescent-shape, positioned transversely; cardiac region longitudinally narrow, wide, positioned transversely, medially separated by shallow setose depression; intestinal region longitudinally narrow, very wide, reaching across most of posterior margin, medially separated by shallow weakly setose part (sometimes not easily discernible); hepatic region obliquely ovate, swollen, smooth, often with median depression, adjacent to external orbital and first anterolateral teeth ( Figs. 2A, B View FIGURE 2 , 3A–C View FIGURE 3 ). Frontal margin slightly produced, gently deflexed, with 2 low lobes separated by shallow concavity; lateral lobe very low, almost undiscernible ( Figs. 3A–C View FIGURE 3 , 5A, B View FIGURE 5 ). Anterolateral margin gently convex, weakly separated from posterolateral margin; external orbital tooth acutely triangular, directed anteriorly, tip sharply pointed to gently rounded; first anterolateral tooth broadly triangular, sharply pointed; second anterolateral tooth smaller than first, tip sharp; 2 large rounded to subtruncate tubercles at junction between antero- and posterolateral margin, tip rounded to flattened, often with median depression ( Figs. 3A–C View FIGURE 3 ). Posterolateral margin gently convex, surface granulated; margin gently converging to sinuous posterior carapace margin ( Fig. 3A–C View FIGURE 3 ). Posterior part of sub-branchial region with distinct rounded ridge, forming channel with margin, connected to end of posterolateral margin, at junction with metabranchial region. Suborbital, subhepatic, pterygostomial and sub-branchial regions smooth ( Fig. 5A, B View FIGURE 5 ). Orbit transverse, large; supraorbital margin gently sinuous, entire; suborbital margin gently convex, entire, with low tooth on inner angle; eyes large with short peduncle and large cornea ( Figs. 3A–C View FIGURE 3 , 5A, B View FIGURE 5 ). Antenna with basal article quadrate, fused with epistome but sutures visible, article 3 slightly shorter than basal article; article 4 as long as article 5, flagellum short, not entering orbit; antennule folding transversely ( Fig. 5A, B View FIGURE 5 ). Epistome wider longitudinally, posterior margin with wide triangular median lobe with median fissure, separated from lateral part by short fissure; lateral margin prominently concave ( Fig. 5A, B View FIGURE 5 ). Endostome with low oblique ridge on each half.
Third maxilliped relatively short; merus quadrate, anteroexternal angle weakly auriculiform; ischium subrectangular with shallow oblique median sulcus; exopod relatively broad, tip not reaching distal edge of merus, with long flagellum ( Fig. 5G, H View FIGURE 5 ).
Chelipeds almost symmetrical ( Fig. 2A, B View FIGURE 2 ), relatively short, covered with same pubescence on carapace except for areolets; basis-ischium lined with small sharp granules on ventral margin; merus short, unarmed, outer surface with 4 transverse tubercles, proximal ones largest; carpus with sharp inner distal tooth, dorsal surface with 1 large oblique tubercle and 2 smaller ones, outer lateral margin with 2 low rounded lobes, basally fused in part, proximal one larger; chela not enlarged, outer surface granulate, fingers longer than palm, dorsal margin of dactylus serrate; propodus not bent downwards, almost subparallel with ventral margin of palm ( Figs. 6A, B View FIGURE 6 , 7A, B View FIGURE 7 ).
Ambulatory legs not prominently elongate, covered with same pubescence on carapace except for swollen surfaces; outer surface weakly eroded; P3 longest, P5 shortest; merus with subdorsal longitudinal swelling, forming low dorsal and ventral cristae; carpus and propodus with prominent median longitudinal swelling, outer surface almost smooth to gently rugose; dactylus gently curved along most of length, tip hooked, glabrous ( Figs. 2A, B View FIGURE 2 , 7G, H View FIGURE 7 ).
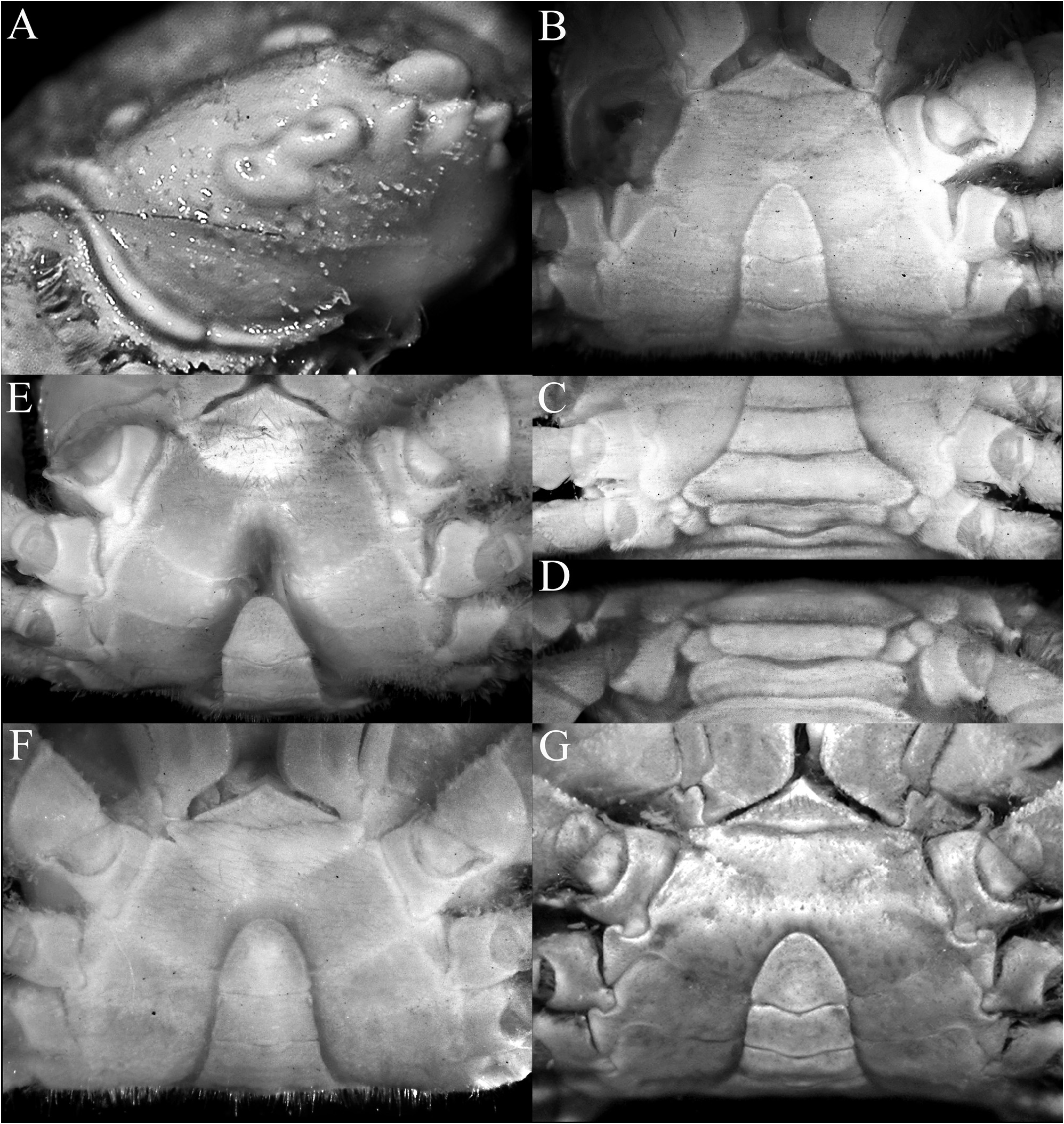
Thoracic sternum relatively smooth, setose; sternites 1 and 2 completely fused to form triangular structure; sternite 2 and 3 separated by distinctly convex suture; sternites 3 and 4 fused but shallow oblique lateral and submedian depression demarcating sternites visible; sternopleonal cavity reaching imaginary line connecting proximal edges of coxae of chelipeds; sternite 8 exposed when pleon closed, visible as subquadrate plate ( Fig. 9B–D View FIGURE 9 ).
Pleon triangular, transversely narrow, especially along somites 4–6; all somites and telson free; somite 1 widest, reaching to P5 coxae; somite 2 less than somites 1 and 3; somite 3 wide, reaching condyles of P5 coxae; somites 4 and 5 trapezoidal with gently concave margins; somite 6 rectangular; telson triangular, slightly longer than wide with rounded tip and gently sinuous lateral margins ( Figs. 9B–D View FIGURE 9 , 10A View FIGURE 10 ).
G1 strongly sinuous, distal part long, gently curved, elongate ( Fig. 10B–D View FIGURE 10 ); G2 short spatuliform ( Fig. 10E View FIGURE 10 ).
Females. Adult female specimens have a longitudinally ovate pleon that covers most of thoracic sternum, telson semicircular ( Fig. 8A View FIGURE 8 ); vulva large, ovate, obliquely positioned, occupying proximal three-quarters of sternite 6, no operculum ( Fig. 8B View FIGURE 8 ).
Colour in life. Background colour of carapace and pereopods white, with brown setae; epigastric areolets red; protogastric areolets bright to pale red; hepatic areolets white to orange; cardiac areolets pale yellow to dirty white ( Fig. 1A, B View FIGURE 1 ; Maenosono 2019: fig. 8A).
Remarks. The taxonomy of L. sculpta has been reviewed by Tesch (1918), Takeda (1977) and Ng (1987). Its original placement in Pilumnoplax Stimpson, 1858 , is not tenable as that genus is now a junior synonym of the euryplacid Eucrate De Haan, 1835 (cf. Castro & Ng 2010). The present specimens agree very well with what was described and figured in Takeda (1977) and Maenosono (2019).
Lophoplax sculpta was described from at least one female specimen (cf. Stimpson 1858: 93; 1907: 91) collected from Ousima, Ryukyu Islands, in Japan, with one measurement provided: 7.1 by 5.6 mm. The specimen is no longer extant (cf. Evans 1967; Deiss & Manning 1981; Manning 1993; Manning & Reed 2006). While the species is distinctive, the descriptions of two new allied species from Indonesia and Vanuatu requires stability in this name, and a neotype is therefore needed. We therefore designate the male specimen examined here as the neotype of Pilumnoplax sculpta Stimpson, 1858 ( Figs. 2A View FIGURE 2 , 3A View FIGURE 3 , 5A View FIGURE 5 , 6A View FIGURE 6 , 7A, G View FIGURE 7 , 9A–D View FIGURE 9 , 10A–E View FIGURE 10 ). It is in good condition and is also from the Ryukyu Islands.
Lophoplax sculpta most closely resembles L. andamanica in carapace features, but as already discussed at length by Trivedi et al. (2022), L. sculpta can easily be separated by the epigastric areolet being more ovate ( Fig. 3A–C View FIGURE 3 ) (versus epigastric areolet quadrilateral in shape in L. andamanica ; Trivedi et al. 2022: fig. 1A); the protogastric areolet is transversely ovate to weakly crescent-shaped in L. sculpta ( Fig. 3A–C View FIGURE 3 ) (versus distinctly crescent-shaped in L. andamanica ; Trivedi et al. 2022: fig. 1A); the hepatic region is proportionately shorter, reaching only to first anterolateral tooth ( Fig. 3A–C View FIGURE 3 ) (versus distinctly longer, reaching to second anterolateral tooth in L. andamanica ; Trivedi et al. 2022: fig. 1A); the tips of the external orbital and anterolateral teeth are sharp ( Fig. 3A–C View FIGURE 3 ) (versus first three anterolateral teeth proportionately broader with the blunt tips in L. andamanica ; Trivedi et al. 2022: fig. 1A); the epistome is proportionately slightly wider ( Fig. 5A, B View FIGURE 5 ) (versus proportionately narrower in L. andamanica ; Trivedi et al. 2022: fig. 1B); the outer surface of the carpus of the cheliped has 2 large tubercles on the dorsal surface and 2 tubercles on the outer margin ( Fig. 6A, B View FIGURE 6 ) (versus outer surface and margin with 6–8 smaller and more dispersed tubercles in L. andamanica ; Trivedi et al. 2022: fig. 1C); the cheliped fingers are not distinctly bent, with the pollex subparallel with the ventral margin of palm ( Fig. 7A, B View FIGURE 7 ) (versus fingers sharply bent downwards in L. andamanica ; Trivedi et al. 2022: fig. 1D); and the outer surface of the P3–P5 merus, carpus and propodus are relatively smoother ( Fig. 7G, H View FIGURE 7 ) (versus surfaces distinctly more eroded in L. andamanica ; Trivedi et al. 2022: fig. 1F–H). It is also useful to note that L. andamanica is only known from the Andamans in the eastern Indian Ocean and is the only species representing the genus from that ocean thus far.
The Philippine specimen (ZRC 2019.1611) differs somewhat from the Japanese specimen and what has been reported by Takeda (1977) and Maenosono (2019) in that the carapace is proportionately slightly wider, the protogastric areolets are weakly crescent-shaped rather than ovate, and the two large tubercles at the junction between the antero- and posterolateral margins are prominently larger ( Fig. 2B View FIGURE 2 , 3C View FIGURE 3 vs. Fig. 2A View FIGURE 2 , 3A View FIGURE 3 ). The two tubercles at the junction between the lateral margins are also slightly larger and not arranged in a linear manner like the Japanese specimens ( Fig. 9A View FIGURE 9 ), appearing as if there are three tubercles rather than two. The differences, however, do not appear significant at the species level and can be explained by intraspecific variation.
For comparisons with L. sordida n. sp. and L. pannosa n. sp., see remarks for these species.
Habitat. From the available data, the species occurs in shallow waters outside reefs. In intertidal areas, it occurs under rocks, often with seagrass nearby. One specimen (ZRC 2019.1611) was collected from a dense seagrass patch with a sandy substrate.
Distribution. Known from the Ryukyus, Japan; and Philippines ( Stimpson 1858; Takeda 1977; Maenosono 2019; Ng et al. 2008).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Lophoplax sculpta ( Stimpson, 1858 )
| Ng, Peter K. L. & Rahayu, Dwi Listyo 2023 |
Lophoplax sculpta
| Maenosono, T. 2019: 30 |
| Castro, P. & Ng, P. K. L. 2010: 25 |
| Naruse, T. 2010: 31 |
| Ng, P. K. L. & D. Guinot & P. J. F. Davie 2008: 143 |
| Sakai, K. 2003: 22 |
| Marumura, M. & Kosaka, A. 2003: 47 |
| Nomura, K. & Nagai, S. & Asakura, A. & Komai, T. 1996: 17 |
| Dai, A. - Y. & Yang, S. - L. 1991: 409 |
| Nagai, S. & Nomura, K. 1988: 196 |
| Ng, P. K. L. 1987: 79 |
| Dai, A. - Y. & Yang, S. - L. & Song, Y. - Z. & Chen, G. - X. 1986: 379 |
| Miyake, S. 1983: 231 |
| Takeda, M. 1977: 120 |
| Sakai, T. 1976: 304 |
| Serene, R. & Lohavanijaya, P. 1973: 69 |
| Serene, R. 1968: 91 |
| Tesch, J. J. 1918: 199 |
Pilumnoplax sculpta
| Stimpson, W. 1907: 91 |
| Miers, E. J. 1886: 226 |
| Stimpson, W. 1858: 93 |