
Philautus kakipanjang, Dehling, J. Maximilian & Dehling, D. Matthias, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3686.2.8 |
|
publication LSID |
lsid:zoobank.org:pub:23C42922-B568-4B62-AC5B-F9E8FED4AEB1 |
|
DOI |
https://doi.org/10.5281/zenodo.5685437 |
|
persistent identifier |
https://treatment.plazi.org/id/039087E2-3176-4124-FF47-724176E2FAA8 |
|
treatment provided by |
Plazi |
|
scientific name |
Philautus kakipanjang |
| status |
sp. nov. |
Philautus kakipanjang View in CoL sp. n.
Western long-legged Bush Frog
Holotype: NMBE 1060433 (ex JMD 323), adult male, from below the summit of Gunung Serapi (01°35.261’ N, 110°11.578’ E; 780 m a.s.l.), Kubah National Park , Sarawak, Malaysia, collected by J. M. Dehling and D. M. Dehling, 25 September 2009 at 0 0 15 h.
Paratypes: NMBE 1060432 (ex JMD 322), adult male, same collection details as for holotype ; NMBE 1060427 – 1060431 (ex JMD 461, 466, 472, 483, 488, respectively), five adult males, from the summit region of Gunung Gading (1°42.813' N, 119°50.186' E; 885 m a.s.l.), collected by J. M. Dehling and D. M. Dehling, 7 October 2009, between 1900 and 2200 h.
Diagnosis: Currently, there are no morphological synapomorphic characters considered diagnostic for the genus Philautus . We assign the new species to this genus for showing morphological and bioacoustic resemblance and close genetic relationship to several of the Bornean species assigned to the genus. The new species differs from its congeners by the combination of the following characters: size moderately large (SVL of adult males 21.2–23.7 mm); hindlimbs long, tibiotarsal joint reaching beyond tip of snout when legs are adpressed to body, TFL/SVL 0.56–0.64; snout acuminate in dorsal view, longer than eye diameter; median lingual process absent; heel without dermal appendage; nuptial pads small, prominent; toes extensively webbed, formula I 2 - -2.25 II 1.5-2.75 III 1.75- 3 IV 2.75-1.5 V; iris golden in life; advertisement call consisting of a single note with 9–12 pulses, lasting 342–478 ms, with a dominant frequency of 2.0–2.7 kHz, moderate frequency modulation and marked amplitude modulation at 18.1–18.5 °C.
Description of holotype: Measurements are provided in Table 1 View TABLE 1 . Body moderately sturdy, widest at temporal region, tapering to groin ( Figure 1 View FIGURE 1 ); head large (HL/SVL 0.36, HW/SVL 0.41), wider than long (HW/HL 1.15); snout long (SL/HL 0.50), acuminate in dorsal view, rounded in profile ( Figure 1 View FIGURE 1 ), projecting beyond lower jaw, wider than long (SL/EE 0.86); canthus rostralis distinct, straight-lined in profile, strongly concave in dorsal view; loreal region oblique, concave; nostrils rounded, directed laterally; situated much closer to tip of snout than to eye (EN/NS 1.45), separated from each other by distance subequal to distance between eye and nostril (NN/EN 0.96); eyes directed anterolaterally, protruding, large (ED/HL 0.45); pupil horizontal; eye diameter slightly shorter than snout (ED/SL 0.91); interorbital distance wider than upper eyelid (IO/EW 1.16) and greater than internarial distance (IO/NN 1.28); tympanum distinctly visible, its posterodorsal fourth concealed under thick layer of skin, separated from eye by one-fifth of its diameter; tympanum diameter two-fifths of eye diameter; upper jaw with dentition; choanae small, oval, located far anterolaterally at margins of roof of the mouth, concealed by upper jaw in ventral view; vomer processes and teeth absent; tongue long (7.9 mm) and broad (4.6 mm at widest point), bilobed for one-fifth of its length, free distally for two-thirds of its length; median lingual process absent; vocal sac single, median, subgular; vocal sac aperture on each side of the mouth, slit-like, long, directed laterally, situated slightly closer to corner of mouth than to base of tongue.
Dorsal surfaces of head, trunk and limbs smooth with several scattered conical tubercles ( Figure 1 View FIGURE 1 ); lateral sides of trunk shagreened; ventral side of head, trunk and thighs areolate, of tibia, tarsus and arms smooth; supratympanic fold thick and conspicuous, slightly curved, extending from posterior edge of eye to level of insertion of forelimb; heel without dermal appendages; vent covered with small dermal flap.
Forelimbs slender; hand moderately large (HND/SVL 0.28); tips of fingers enlarged into broad oval disks, those of Fingers III and IV twice as wide as penultimate phalanges ( Figure 2 View FIGURE 2 ); each disc with circummarginal groove; relative length of fingers: I <II <IV <III; subarticular tubercles rounded, well developed, numbering one on Fingers I and II, two on Fingers III and IV, with proximal tubercles on Fingers III and IV much smaller than distal ones; rudimentary webbing between fingers, formula being I 2.5-2.75 II 2.5-3.75 III 3 - -2.5 IV ( Figure 2 View FIGURE 2 ); thenar tubercle distinct, about one-fourth size of thenar; palmar tubercles absent; nuptial pad on dorsal surface of Fingers I, II and III, most conspicuously developed as prominent, small (about the size of disc of Finger I), smooth pad on proximal phalanx of Finger I, otherwise as callous tissue on penultimate phalanx of Finger I and penultimate and proximal phalanx of Finger II, and as callous, scattered tubercles on Finger III.
Hindlimbs slender, long (LEG/SVL 1.72); tibiotarsal articulation reaching about one ED beyond tip of snout when legs are adpressed forwardly to body; tibiofibula long (TFL/SVL 0.56), longer than thigh (TFL/THL 1.06); heels overlapping each other slightly when knees flexed and thighs held perpendicularly to median plane; foot shorter than tibiofibula (FOT/TFL 0.79); tips of toes enlarged into broad oval disks, each with circummarginal groove; discs of toes smaller than those of fingers; relative length of toes: I <II <III <V <IV; subarticular tubercles numbering one on Toes I and II, two on Toes III and V, and three on Toe IV; pedal webbing formula I 2 - - 2.25 II 1.5-2.75 III 1.75- 3 IV 2.75-1.5 V ( Figure 2 View FIGURE 2 ); inner metatarsal tubercle relatively large (length 1.0 mm), onethird of length of Toe I (3.0 mm), elongated, prominent; outer metatarsal tubercle absent.
Colouration: Dorsal colouration in life varied strikingly between night and day ( Figure 1 View FIGURE 1 ). During the night, dorsum reddish-cream-coloured with faint greyish-brown markings; light brown stripes along the canthus rostralis, between the eyes, and parallel to spine on dorsum; limbs with faint crossbars. During the day, dorsum light brown with dark brown, conspicuous markings; sides of the head below canthus rostralis, eye, and supratympanic fold almost uniformly dark brown; tympanum yellowish brown; interorbital stripe, crossbars on limbs, and stripes on the back darker and wider than during the night; additional dark brown flecks on dorsum visible. Chin, throat, chest, and ventral surfaces of forelimbs whitish, speckled with yellow, flanks yellow. Venter and ventral surfaces of hindlimbs mostly unpigmented. Preaxial side of thigh and groin weakly speckled with yellow markings. Iris golden. In preservative, pattern and colouration similar to diurnal colouration.
Variation: The paratypes are very similar to the holotype in measurements and proportions ( Table 1 View TABLE 1 ). All paratypes have a median subgular vocal sac and small, smooth, low nuptial pads. Dorsal pattern varies considerably ( Figure 1 View FIGURE 1 ). One paratype (NMBE 1060429) has a broad longitudinal mid-dorsal band, another one (NMBE 1060431) a fine mid-dorsal stripe. Mottling on the ventral side and yellow colouration on thigh is variable. So far, females of this species are unknown.
Molecular genetics: Comparison of a fragment of the 16S rRNA sequence showed that the species differs considerably from all other species of the genus for which homologous information is available. The new species differs in the uncorrected p-distance from the most closely related Bornean and Philippine species by 9.9–13.4 % (Table 2). Between the comparative species examined, p-distances ranged between 3.5 and 16.0 % (Table 2).
Advertisement call: 34 advertisement calls from six individuals including the holotype and three of the paratypes were recorded in the field (air temperature was 18.5 °C at Gunung Serapi and 18.1°C at Gunung Gading). The advertisement call consists of a single note composed of 9–12 (median 11) pulses and lasting 342– 349 ms (nine pulses), 377–383 ms (ten pulses), 420–425 ms (eleven pulses) or 462–478 ms (twelve pulses). Individual pulses had a length of 4 ms. In one-third of the calls, the note was followed by one or two additional pulses after an interval of 155–220 ms. Pulse repetition rate in the main note ranged between 22.7/s and 26.3/s (mean 24.7/s). In about half of the calls, pulse repetition rate at the beginning of the call was lower than at the end of the note. In the other half of the calls, pulse repetition rate was consistent within the note. In the three cases in which two pulses were added to the note after an interval, the additional two pulses were emitted at a rate of 25.0, 27.0, and 35.7/s. Calls were amplitude-modulated. Amplitude of the pulses was lowest at the beginning of the note, reached maximum at about the middle of the call and remained at that level until the end of the call ( Figure 3 View FIGURE 3 ). The note was slightly frequency-modulated and the first two or three pulses had a lower dominant frequency (energy maximum at 2180 ± 150 [2000–2500] Hz) than the rest of the pulses (energy maximum at 2518 ± 121 [2300–2700] Hz). Increase of frequency within a note was 339 ± 200 (100–700) Hz. When additional pulses were added, their frequency was as high as the frequency of the pulses at the end of the main note.
Habitat and distribution: We found all specimens after dusk, calling from low bushes and from tops of small trees, ca. 0.5–3 m above the ground. We heard the call of the species only in the summit regions of both Gunung Serapi and Gunung Gading from approximately 750 m upwards, where the lowland mixed dipterocarp forest is replaced by the lower montane forest ( Figure 4 View FIGURE 4 ). We recorded calls assignable to the new species at a similar elevation on Gunung Santubong ( Figure 4 View FIGURE 4 ) but were not able to climb to the calling specimens. The species is probably more widespread in the mountains of western Sarawak or even the whole of Borneo.
Etymology: The species epithet is composed of the two Malay words kaki, meaning leg, and panjang, meaning long; in reference to the relatively long hind limbs of the species. The composed word is used as a noun in apposition.
Comparisons: Morphologically, Philautus kakipanjang is most similar to species of the P. aurifasciatus (Schlegel) group sensu Dring (1987; see also Matsui 2009, Dehling 2010) which are all small-sized, have the musculus cutaneus pectoris and small, smooth nuptial pads (in males), and lack vomerine teeth. It is compared in the following paragraph to all other species that share this combination of characters. All other Philautus species of the Sunda region lack this character combination and therefore can be distinguished morphologically from Philautus kakipanjang .
In the following, P. kakipanjang (characters in parentheses) is first compared to Bornean species, then to species with a type locality outside Borneo. Philautus amoenus Smith has shorter legs, with TFL/SVL 0.45–0.54 in males (0.56–0.64) and the tibiotarsal articulation reaching to tip of snout or slightly beyond (reaching considerably beyond tip of snout); a tubercle present at the tibiotarsal joint (absent); a wider head, HW/SVL 0.40–0.47 (0.38– 0.42); and Toes III and V reaching distinctly beyond the knee when tarsus is adpressed to tibia (toe tips reaching to knee only). Philautus davidlabangi Matsui has a longer head, HL/SVL 0.40–0.43 (0.32–0.37); shorter legs, with TFL/SVL 0.52–0.56 (0.56–0.64), tibiotarsal articulation reaching to tip of snout only (reaching considerably beyond tip of snout) and heels meeting or at most slightly overlapping each other (heels overlapping considerably); snout shorter than the eye diameter (longer than eye diameter); interorbital distance shorter than upper eyelid width with IO/EW 0.78–0.90 (greater than EW with IO/EW 1.02–1.21); more extensive toe webbing with I (1.5-1.75)- 2.25 II 1 -2.25 III 1- 2 IV 2– 1 V (vs. I 2 - -2.25 II 1.5-2.75 III 1.75- 3 IV 2.75-1.5 V); and nuptial pads absent (present). Philautus juliandringi Dehling has a shorter snout, subequal in length to eye diameter, SL/ED 0.92–1.05 (longer, SL/ED 1.10–1.25) and rounded in dorsal view (acuminate); a relatively larger eye, ED/HL 0.46–0.52 (0.42–0.45); the tympanum is smaller, TD/ED 0.32–0.39 (0.39–0.48) and is covered by a thick layer of skin (conspicuously visible, free except posterodorsal fourth); and the toe webbing is a little less extensive, I 2 -2.5 II (1.5–2-)-3 III 2 - - 3+ IV (3+–3)- 2 V (vs. I 2 - -2.25 II 1.5-2.75 III 1.75- 3 IV 2.75-1.5 V). Philautus mjobergi Smith has a rounded snout in dorsal view (acuminate), which is shorter or subequal to eye diameter (longer than eye diameter); a relatively wider head, HW/HL 1.29–1.34 (1.14–1.21); the tympanum is separated from the eye by a distance equal to TD (almost immediately adjacent to eye, separated by less than one-fifth of TD); and the toe webbing is less extensive, Toes II, III, and V not webbed beyond distal, Toe IV not beyond penultimate subarticular tubercles (toes webbed beyond these tubercles). Philautus petersi Boulenger has shorter legs with the tibiotarsal articulation reaching to tip of snout only (reaches beyond tip of snout); a median lingual process (absent); a conical tubercle at the tibiotarsal joint (absent) and the tympanum is separated from the eye by two-thirds the TD (almost immediately adjacent to eye, separated by less than one-fifth of TD). Philautus refugii Inger and Stuebing is smaller, SVL of males 15.3– 18.0 mm (21.2–23.7 mm), has less extensively webbed toes, webbing not reaching beyond distal subarticular tubercles of Toes III and V (reaching beyond these tubercles); and a conical tubercle at the tibiotarsal joint, followed by a row of lower tubercles along the postaxial edge of the tarsus and fifth toe (absent); and the anterior face of the thigh has a barred black and white pattern (unpigmented with bright yellow spots). Philautus saueri Malkmus and Riede has shorter legs, TFL/SVL 0.54 (0.56–0.64); a wider head, HW/SVL 0.44 (0.39–0.42); a larger interorbital distance with IO/EW 1.90 (0.97–1.21); the nostril situated between the eye and the tip of snout, EN/NS 1.01 (1.45–1.59); and a small tubercle at the tibiotarsal joint (absent). Philautus umbra Dring is larger, SVL of males up to 35.1 mm (to 23.7 mm); has a median lingual process (absent); less developed toe webbing; no finger webbing (rudimentary webbing); broader finger and toe tips; colouration light grey with darker markings during the night and almost uniformly black during the day (light brown to yellowish brown with or without marked dark brown pattern during the night, light brown with darker markings during the day); and has the nuptial pads reduced to glandular patches (nuptial pads small, prominent, rounded). Philautus acutirostris (Peters) (including its synonyms P. basilanensis Taylor and P. woodi Stejneger ) from the Philippines has a sharply pointed snout (acuminate), frequently with a pale-coloured projection at tip (absent at least in males); less extensive toe webbing, Toe IV webbed only to proximal subarticular tubercle (webbed beyond middle tubercle on postaxial side); a median lingual process (absent); and can have vomerine teeth (absent). Philautus aurifasciatus from Java has shorter legs, TFL/SVL 0.49–0.58 (0.56–0.64), the tibiotarsal articulation reaching to nostril or the tip of snout (reaching beyond tip of snout); and has an NN/EN ratio of 1.11–1.32 (0.87–0.96). Philautus leitensis (Boulenger) from the Philippines has a relatively narrower head with HW/HL 1.03–1.17 (1.14–1.25); an indistinct tympanum (distinct); a prominent tubercle at the tibiotarsal articulation (absent); and lacks finger webbing (rudimentary webbing present). Philautus longicrus Boulenger has a relatively narrower head, HW/HL 1.08–1.16 (1.14–1.21); the internarial distance is larger than the distance from eye to nostril NN/EN 1.09–1.26 (NN shorter than EN, 0.87– 0.96) and larger in relation to head width, NN/HW 0.26–0.29 (0.23–0.26); the nostril is situated a little closer to the tip of snout than to the eye, EN/NS 1.07–1.18 (much closer to tip of snout than to eye, EN/NS 1.45–1.61); the interorbital distance is larger in relation to the width of the upper eyelid IO/EW 1.25–1.34 (0.96–1.21); and the hand is relatively shorter HND/SVL 0.26–0.29 (0.29–0.32). Philautus schmackeri (Boettger) (including P. mindorensis [Boulenger]) from the Philippines has a relatively narrower head with HW/HL 1.00–1.11 (1.14–1.25); a smaller tympanum with TD/ED 0.20–0.25 (0.39–0.48); and a pointed snout with a conical tip (acuminate without conical tip). Philautus parvulus (Boulenger) from Myanmar, Thailand, Cambodia, Vietnam, and Peninsular Malaysia has shorter legs with TFL/SVL 0.44–0.51 (0.56–0.64) and the tibiotarsal articulation reaches the posterior margin of eye (beyond tip of snout); the heels barely touch when legs are folded at a right angle to the body (overlapping considerably).
Bioacoustically, Philautus kakipanjang is also most similar to the Bornean species of the P. aurifasciatus group sensu Dring (1987; see also Matsui 2009, Dehling 2010) and is compared to these species in the following paragraph. Calls of these species are regularly pulsed and not or only slightly frequency-modulated. All other Bornean Philautus species, except P. disgregus Inger and P. erythrophthalmus Stuebing and Wong , whose calls are unknown, can be distinguished from P. kakipanjang by their strongly differing advertisement calls ( Dring 1987, Malkmus & Riede 1996, Malkmus et al. 2002, Dehling 2010, Dehling et al. 2010, unpubl. data of JMD). The calls of the following species differ in main characteristics (in parentheses: number of notes, note length, number of pulses, dominant frequency, ambient temperature, respectively), and thus can be used to distinguish these species from P. kakipanjang (1 note, 342–478 ms, 9–12 pulses, 2.0–2.7 kHz, 18.1–18.5°C): P. amoenus (2–3 notes, 110 ms, 3–4 pulses, 1.8 kHz, 12.5°C; Malkmus & Riede 1996, Malkmus et al. 2002); P. davidlabangi (1 note, 50–80 ms, 13–18 pulses, 2.9–3.3 kHz, 23.5–24.8°C; Matsui 2009, unpubl. data of JMD); P. juliandringi (1 note, 234–658 ms, 2–9 pulses, 2.6–3.5 kHz, 18.3–19.1°C; Dehling 2010); P. mjobergi (call series of 1–4 calls, calls consisting of 1–11 notes, 20–70 ms, 2–7 pulses, 2.0–2.8 kHz, 15 °C; Dring 1987); P. petersi (1 note, 1180–1300 ms, 51–56 pulses, 1.6–2.2 kHz, 18°C; Dring 1987); P. refugii (1–3 notes, 10–20 ms, 2–5 pulses, 3.5–4.0 kHz, 18.1°C; Dehling & Dehling, unpubl.); P. saueri (2–4[rarely 5] notes, 30–40 ms, 7–8 pulses, 1.9–2.6 kHz, 15°C; Malkmus & Riede 1996, Malkmus et al. 2002); P. u m b r a (1 note, 120–587 ms, 2–11 pulses, 1.4–2.1 kHz, 18.3–19.1°C; unpubl. data of JMD). The call of P. parvulus from Thailand differs in consisting of 1–7 single-pulsed notes, each lasting 20–40 ms at unknown temperature ( Heyer 1971).
Finally, the partial 16S rRNA sequence of Philautus kakipanjang differs by at least 9.9 % in the uncorrected pdistance from all available sequences of other species of the genus, including the species which are morphologically and bioacoustically most similar. This value is much higher than the distances between morphologically and bioacoustically well distinguished species pairs like P. acutus Dring / P. aurantium Inger (3.5 %), P. aurantium / P. bunitus Inger, Stuebing and Tan (4.8 %), or P. everetti Boulenger / P. macroscelis Boulenger (5.0 %) (Table 2), corroborating the distinct specific status of P. kakipanjang .

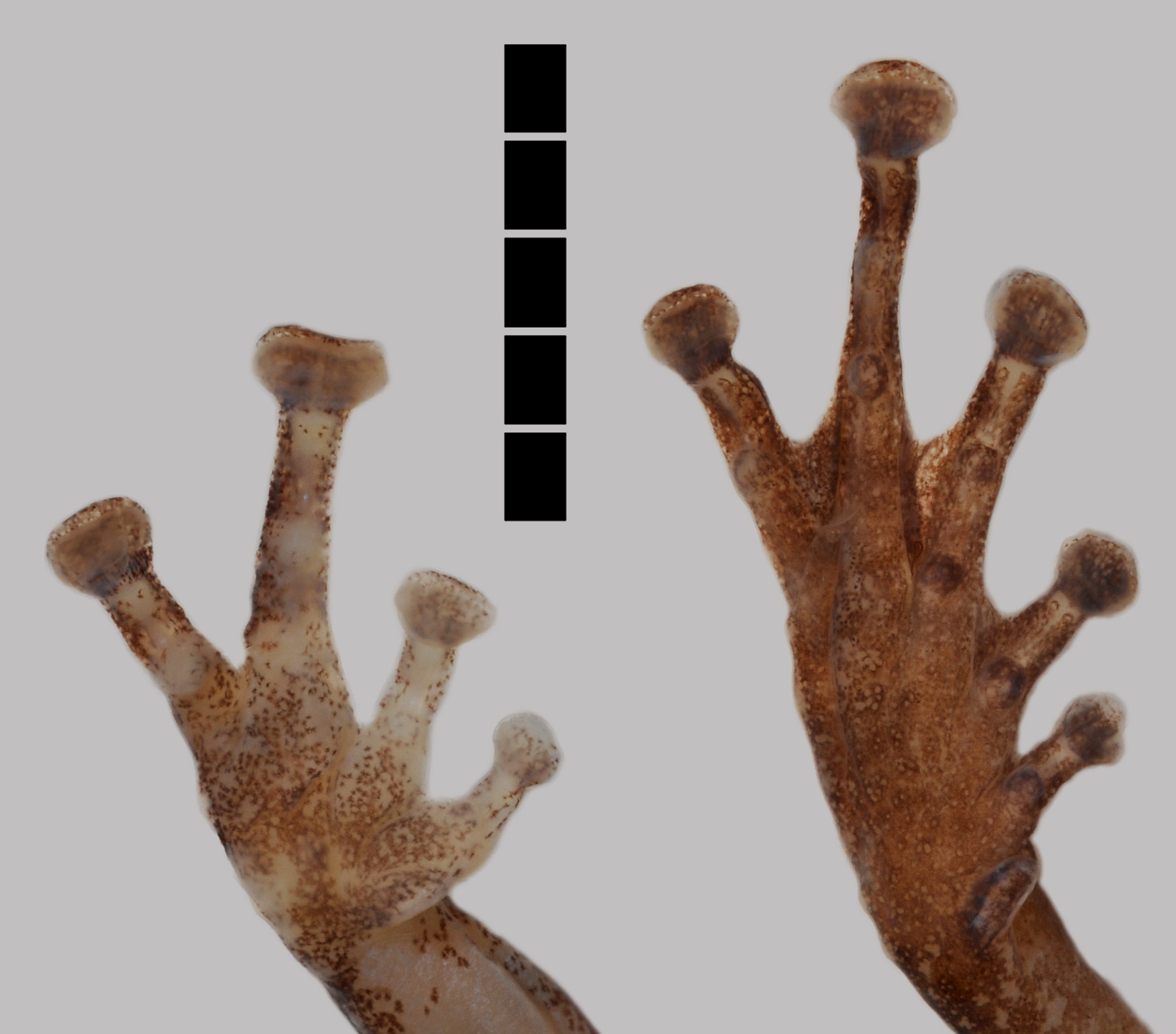

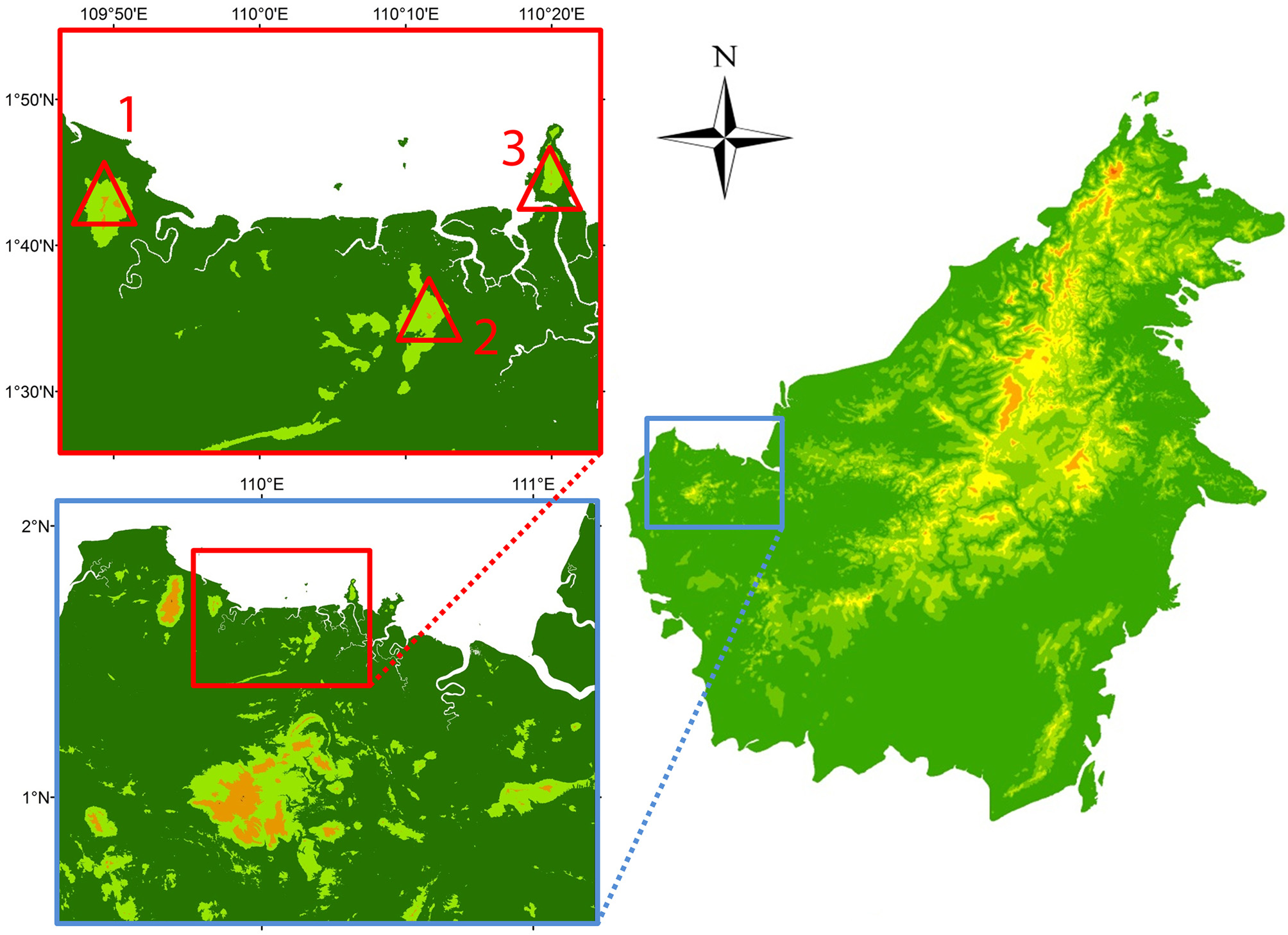
TABLE 1. Measurements of the type series of Philautus kakipanjang sp. n. For abbreviations see Material & Methods.
| NMBE | 1060433 | 1060432 | 1060427 | 1060428 | 1060429 | 1060430 | 1060431 |
|---|---|---|---|---|---|---|---|
| status | holotype | paratype | paratype | paratype | paratype | paratype | paratype |
| origin | Kubah NP | Kubah NP | Gading NP | Gading NP | Gading NP | Gading NP | Gading NP |
| SVL | 21.4 | 22.0 | 21.4 | 22.3 | 23.7 | 21.4 | 21.2 |
| TFL | 12.1 | 12.7 | 12.7 | 13.7 | 13.6 | 12.6 | 13.6 |
| FOT | 9.5 | 9.6 | 9.4 | 10.8 | 10.5 | 9.9 | 10.3 |
| TarL | 15.1 | 15.8 | 15.8 | 16.8 | 17.2 | 16.2 | 16.7 |
| LEG | 36.8 | 37.4 | 37.3 | 41.0 | 41.5 | 38.4 | 40.6 |
| THL | 11.4 | 11.7 | 11.4 | 12.7 | 12.6 | 12.2 | 12.4 |
| ARM | 10.7 | 11.0 | 10.8 | 11.4 | 12.5 | 10.9 | 11.1 |
| HND | 6.2 | 6.3 | 6.5 | 7.0 | 7.6 | 6.9 | 6.7 |
| HW | 8.8 | 9.3 | 8.3 | 8.9 | 9.7 | 8.5 | 8.7 |
| HL | 7.6 | 8.0 | 6.9 | 7.4 | 8.5 | 7.4 | 7.2 |
| IO | 2.8 | 2.7 | 2.5 | 2.4 | 2.8 | 2.4 | 2.7 |
| EW | 2.4 | 2.6 | 2.1 | 2.2 | 2.4 | 2.1 | 2.2 |
| ED | 3.4 | 3.6 | 2.9 | 3.2 | 3.7 | 3.1 | 3.2 |
| TD | 1.3 | 1.4 | 1.4 | 1.3 | 1.4 | 1.4 | 1.4 |
| ET | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 |
| EN | 2.8 | 2.5 | 2.3 | 2.4 | 2.7 | 2.2 | 2.3 |
| EE | 4.4 | 4.5 | 4.2 | 4.3 | 4.9 | 4.1 | 4.2 |
| NS | 1.6 | 1.5 | 1.5 | 1.5 | 1.7 | 1.5 | 1.5 |
| SL | 3.8 | 4.0 | 3.7 | 3.8 | 4.3 | 3.6 | 3.6 |
| NN | 2.2 | 2.3 | 2.1 | 2.2 | 2.5 | 2.1 | 2.0 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.