
Bruaylogus magnificus, Coty, David, Háva, Jiří, Prokop, Jakub, Roques, Patrick & Nel, André, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3878.5.4 |
|
publication LSID |
lsid:zoobank.org:pub:377F9037-D039-44A5-B562-051560CBC9E0 |
|
DOI |
https://doi.org/10.5281/zenodo.6142665 |
|
persistent identifier |
https://treatment.plazi.org/id/03923C21-8625-FF89-FF45-F9510541F9D6 |
|
treatment provided by |
Plazi |
|
scientific name |
Bruaylogus magnificus |
| status |
sp. nov. |
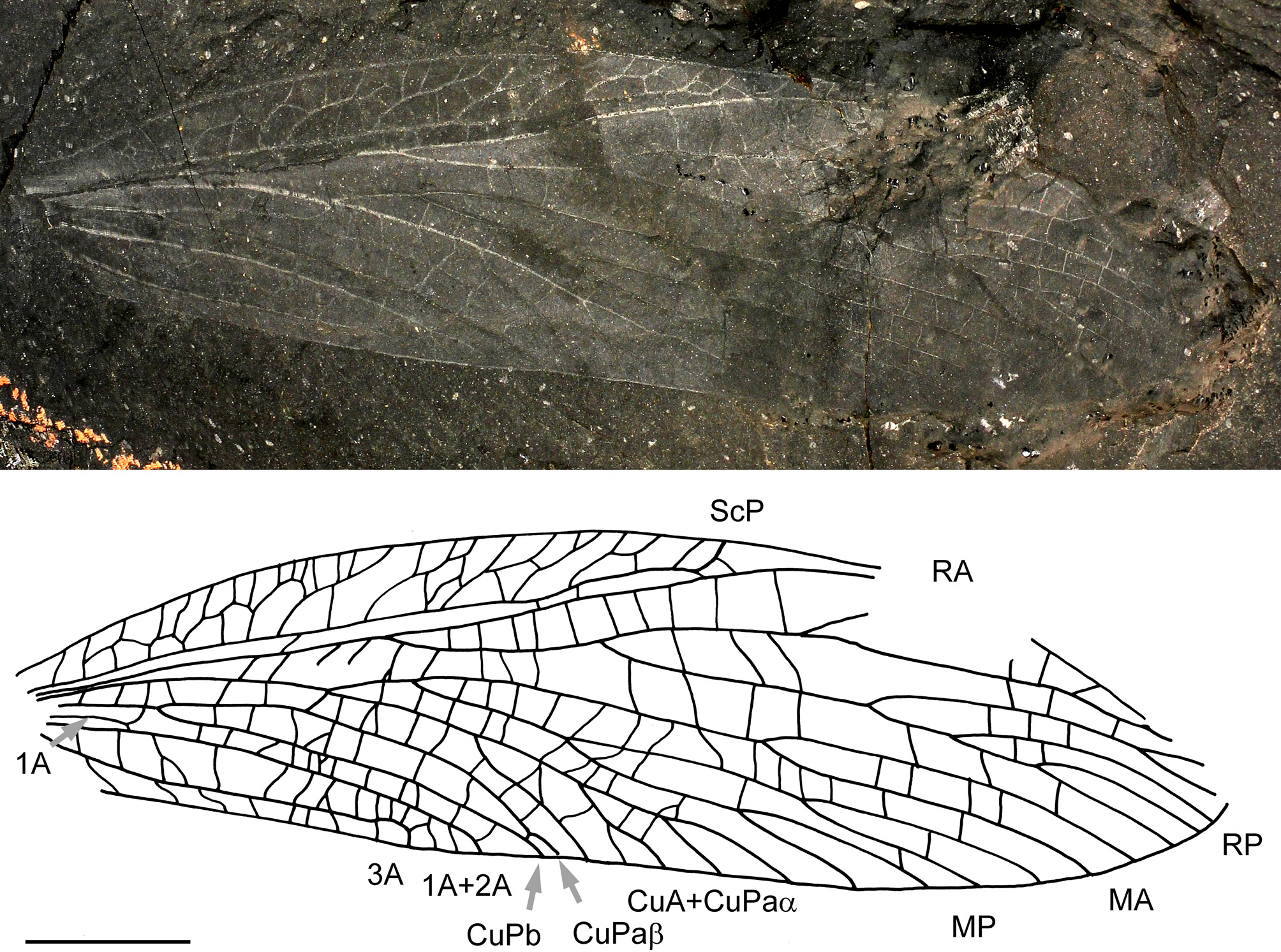
Bruaylogus magnificus sp. nov.
( Fig. 2 View FIGURE 2 )
Material studied: Holotype specimen Bruay 00058a (counterpart), coll. Patrick Roques, to be deposited in the Musée Géologique Pierre Vetter, Decazeville, France.
Age and outcrop. Upper Carboniferous, Bashkirian, Westphalian C or D, Bruay-en-Artois, coal heap n° 26, Department of North, France.
Etymology. Named after the wonderful state of preservation of the fossil.
Diagnosis. As for the genus.
Description. Counterpart of a nearly complete forewing perfectly preserved with only anterior part of apex missing. Preserved length 37.5 mm, width 10.1 mm; no defined ScA; ScP anteriorly pectinate, ending on anterior wing margin, with long branches forked and sigmoidal; area between costal margin and ScP 2.0 mm wide; base of RP 11.5 mm distal of wing base; ScP and R (+ M + CuA) basally separated; presence of a common stem R + (M + CuA); RA convex, simple; area between RA and RP with straight cross-veins; RP rather concave; first fork of RP 6.8 mm from its base, anterior branch with at least one more fork distally, posterior branch anteriorly pectinate; CuA separated from M in basal third of wing; M dichotomously ramified into neutral MA and concave MP; MA well distant from RP, with only one distal fork; MP dichotomously ramified two times (four branches reaching posterior wing margin); CuA fused with CuPaα at base of M; CuA + CuPaα posteriorly pectinate, with its two first posterior branches concave; CuPaα short basal of its fusion with CuA, 0.8 mm long; CuPaβ simple; a series of short crossveins in area between CuA + CuPaα and CuPaβ; division of CuP into CuPa and CuPb 4.0 mm from wing base; CuPb simple; area between CuPa and CuPb narrow and narrowed near posterior wing margin (largest width opposite origin of CuPaα / CuPaβ); three anal veins, all simple; anterior one 1A very short and fused to 2A well separated from CuP at wing base.
Discussion. This forewing corresponds to that of a clade Panorthoptera Crampton 1928 sensu Béthoux & Nel (2002: 27) because of the following synapomorphies: CuPa differentiated into two branches (CuPaα and CuPaβ) just basal of the fusion of the anterior one (CuPaα) with CuA (forewing autapomorphy of Panorthoptera); ScP reaching anterior wing margin and not RA; MA1 and MA2 can be differentiated; CuA + CuPaα ramified with at least three branches; CuPb simple. Bruaylogus gen. nov. shares with the Orthoptera Olivier, 1789 (sensu Béthoux & Nel, 2002: 38-39) the MA simply forked into MA1 and MA2; and CuPaβ, CuPb and 1A simple. But its MP is divided into four distal branches instead of being simple or with few distal branches and it is not especially very concave. Thus if it certainly belongs to the stem lineage of the Orthoptera , it cannot be included in the crown group of this clade.
A pattern of venation rather similar to that of Bruaylogus can be found in the Middle Permian Permostridulidae Béthoux et al., 2003 [a family of the panorthopteran order Caloneurodea Handlirsch, 1937 (sensu Béthoux et al., 2004)]: presence of a forked MA, MP with two dichotomies, and CuPaβ, CuPb and 1A simple. The crucial differences between Bruaylogus and Permostridulus Béthoux et al., 2003 are as follows: the vein CuA + CuPaα posteriorly pectinate in the former while it is simple in the latter taxon; CuPaβ and CuA + CuPaα strongly approximate in Permostridulus (apomorphy of the Caloneurodea), while they are less close in Bruaylogus (Béthoux et al., 2003).
The Oedischiidae Handlirsch, 1906 View in CoL , probably the most basal lineage of the Orthoptera View in CoL (see Béthoux & Nel, 2002), differ from Bruaylogus in the veins RP and MA1 strongly approximate or fused for a short distance and the vein MP simple or with weak distal branches ( Carpenter, 1992; Beckemeyer, 2011). Nevertheless Bruaylogus shares with the Oedischiidae View in CoL the regular space between the posterior branches of CuA + CuPaα and basal fork of M rather close to point of separation between M and CuA. The Geraridae Scudder, 1885 is a panorthopteran group whose monophyly and affinities with the Orthoptera View in CoL sensu stricto are not clear (see Béthoux & Nel, 2002). All the taxa currently included in this family (see Zessin, 2012) differ from Bruaylogus in the three characters: RA with well defined branches, MA (or its anterior branch) and RP strongly approximate or fused for a short distance, CuPb with numerous branches (this last character is unknown in Hirsutgerarus Zessin, 2012 and Progenentomum Handlirsch, 1906) ( Burnham, 1983; Kukalová-Peck & Brauckmann, 1992; Béthoux & Nel, 2002, 2003; Zessin, 2009, 2012). Note that Zessin (2012) listed Anepitedius giraffa Handlirsch, 1911 in the gerarids, but Béthoux & Nel (2002: 32) considered it as a ‘Protorthoptera’ incertae sedis, which means that it is a very poorly known taxon. Lastly Heterologus duyiwuer Béthoux et al., 2012 and Heterologus langfordorum Carpenter, 1944 share with Bruaylogus the median and radial veins well separated, a simple CuPaβ, and M with numerous branches, but both strongly differ from Bruaylogus in the long stem of M before its first fork ( Carpenter, 1944; Béthoux et al., 2012). Furthemore, H. duyiwuer differs from Bruaylogus in the distally forked CuPb and first branch of CuA + CuPaα in a basal position. A further difference between Heterologus duyiwuer and Heterologus langfordorum is the presence of three anal veins reaching posterior wing margin in H. duyiwuer while there are only two in H. langfordorum (after the photograph of the type in Carpenter, 1944: pl. 3), as in Bruaylogus . Furthermore Bruaylogus and H. langfordorum share the second visible anal vein (3A) very long and parallel with posterior wing margin. Possibly H. langfordorum has the first anal vein 1A fused with second 2A as in Bruaylogus .
In conclusion Bruaylogus differs from all described Panorthoptera nec Orthoptera View in CoL in a series of characters that justify a new genus and species. Bruaylogus could be closer to Orthoptera View in CoL in some characters (pattern of stem of M, regular branching of CuA + CuPaα) but the character “ RP and M not approximating” suggests a more basal position for Bruaylogus than the Geraridae , maybe close to the Heterologus spp. A new phylogenetic analysis of the basal taxa of the Panorthoptera becomes necessary after the recent discoveries of the last 10 years.
Remark. Another wing fragment (specimen 00058b, see figure 3) is fossilised 35 mm from the specimen 00058a. It corresponds to the apical part of a wing attributable to a Polyneoptera, and possibly to an Archaeorthoptera. It is probably not the same taxon as specimen 00058a because the distribution and shape of the crossveins are different (denser and more irregular pattern in specimen 00058b).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Bruaylogus magnificus
| Coty, David, Háva, Jiří, Prokop, Jakub, Roques, Patrick & Nel, André 2014 |
Oedischiidae
| Handlirsch 1906 |
Geraridae
| Scudder 1885 |