
Entomobrya silvicola Winkler, Grujić & Fiera, 2023
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5352.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:1825DEE0-348B-44BF-A1DD-26974C269EA3 |
|
DOI |
https://doi.org/10.5281/zenodo.8431660 |
|
persistent identifier |
https://treatment.plazi.org/id/039287B1-FA29-FFB5-FF2A-FA5BFEAEFD68 |
|
treatment provided by |
Plazi |
|
scientific name |
Entomobrya silvicola Winkler, Grujić & Fiera |
| status |
sp. nov. |
Entomobrya silvicola Winkler, Grujić & Fiera sp. nov.
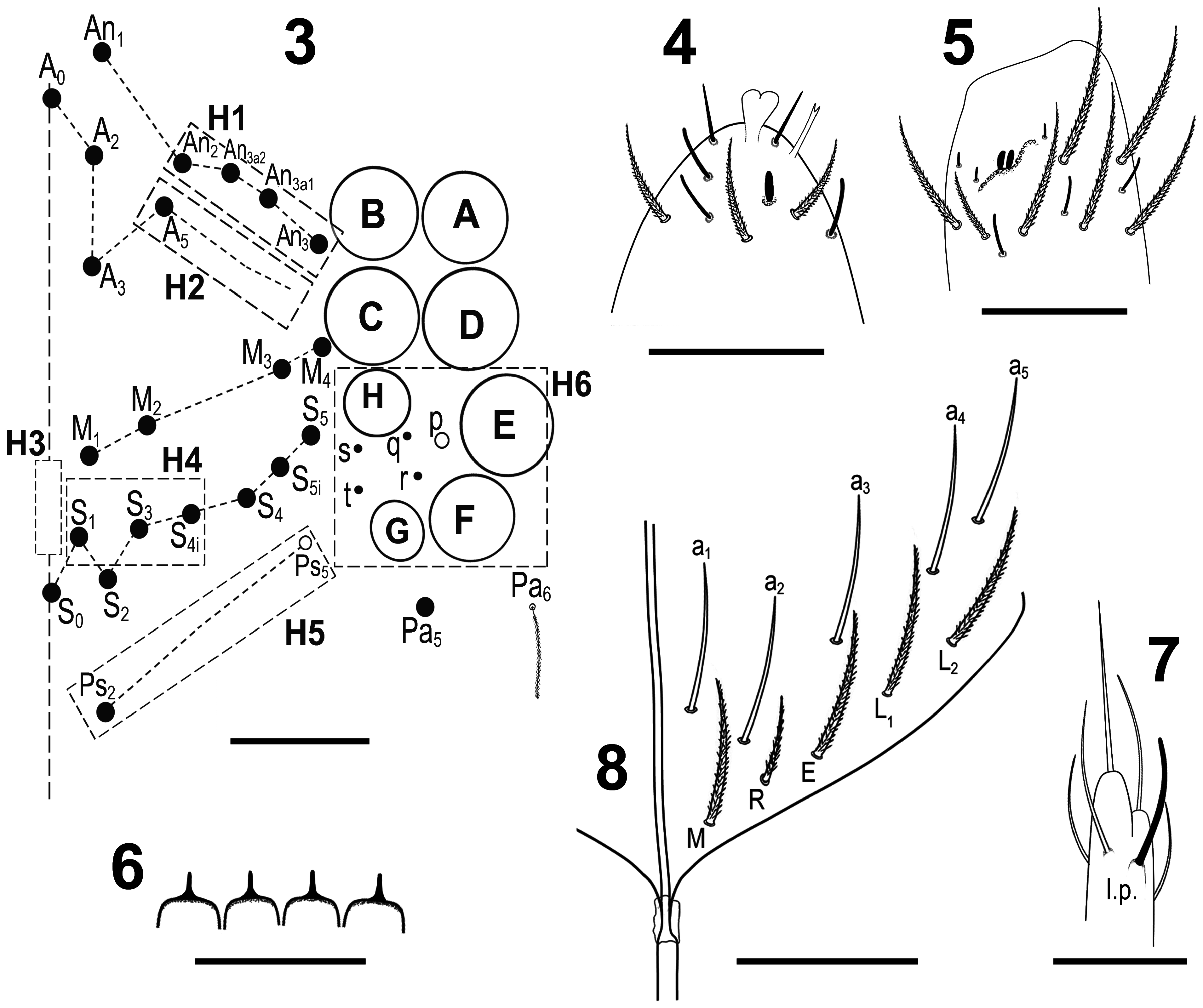
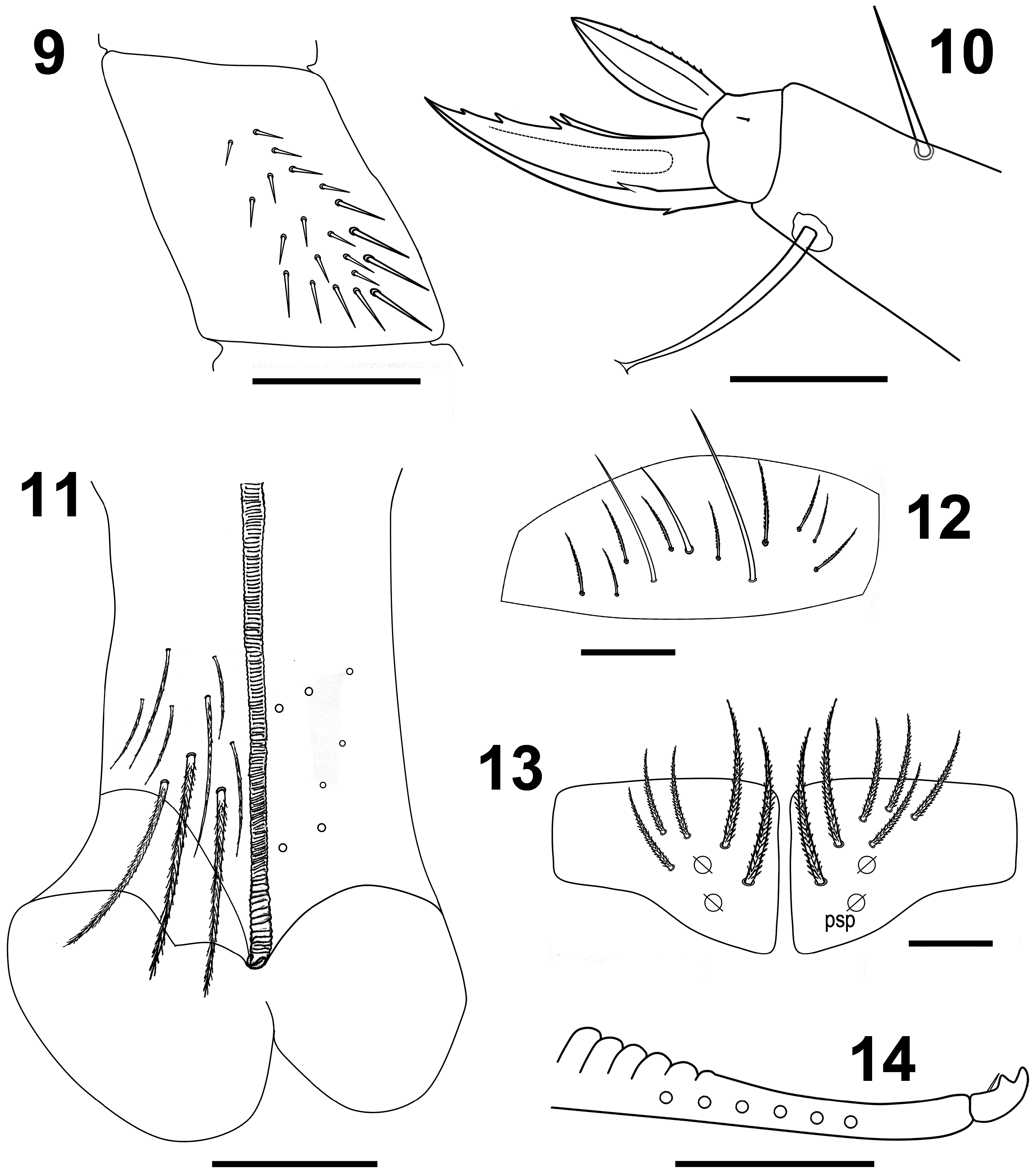
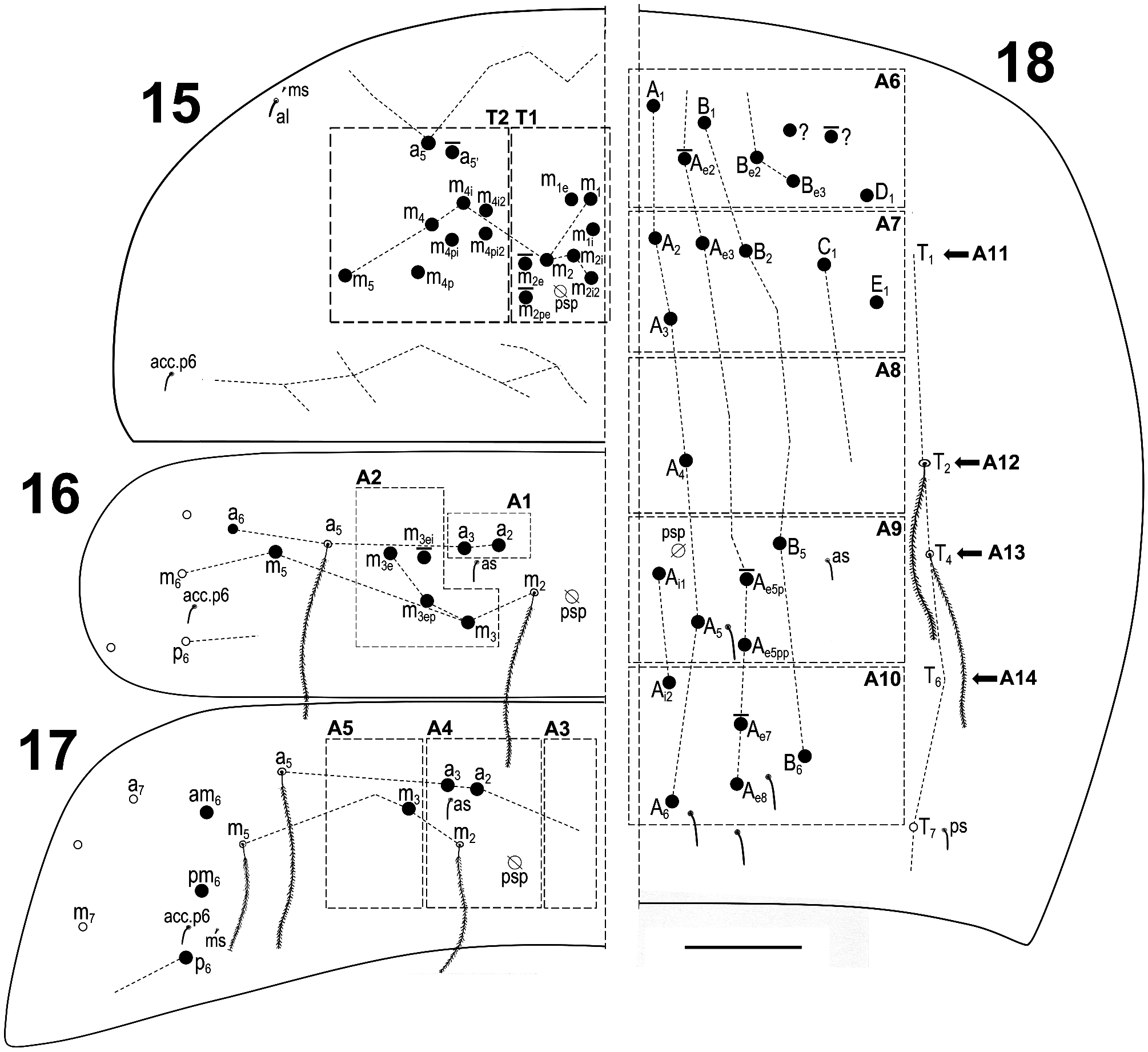
Figs 2–18 View FIGURE 2 View FIGURE 3–8 View FIGURE 9–14 View FIGURE 15–18 , Table 1 View TABLE 1
Type material. Holotype: female on slide (slide code: HNHM-collpr-904), Baške Oštarije ( Croatia), 957 m a.s.l., N 44°32’7”; E 15°8’30”, from forest litter, hand collecting, 20.viii.2022, leg. D. Winkler. GoogleMaps Paratypes (same data as holotype): female on slide (slide code: HNHM-collpr-905) and one juvenile on the same slide as the holotype. The holotype and GoogleMaps paratypes are deposited in the Hungarian Natural History Museum ( HNHM), Budapest GoogleMaps .
Diagnosis. Body pale yellow with purple-brown transverse stripes continuous on Abd II–III and centrally interrupted on Th II–Abd I, but continually broadened laterally on posterior margins. Abd IV with irregular lateral patches. Ant IV with bilobed apical bulb. Labral papillae with spine-like projection. Lateral process on labial papilla E outreaching apex of papilla. Claw with four inner teeth, paired lateral teeth intermediate, unpaired dorsal tooth basal. The exact identification of the species can be made by using the abbreviated macrochaetotaxy formula (sensu Jordana & Baquero 2005) of head (H1–5 areas), Th II (T1–2 areas), Abd II (A1–2 areas), Abd III (A3–5 areas), and Abd IV (A6–10 areas) as:
4-1-0-3-1/6(8)-8(9)/2-3(4)/0-2-1/6(8)-6-1-4(5)-4(5).
Description. Habitus. Adult body length (excluding antennae) 2.22–2.56 mm (n=2), holotype 2.22 mm. Adult body ground pale yellow ( Figs 2A–B View FIGURE 2 ). Pattern with purple-brown transverse stripes on posterior margins of Th II–Abd III, continuous on Abd II–III and centrally interrupted on Th II–Abd I, but always broadened laterally. Abd IV with irregular purple-brown lateral patches. Dorsal head with dark spot between bases of antennae and broad purple-brown shade along centreline and lateroventral area. Dark violet pigment on antennae with increasing intensity from base to apex of segments. Dark purple-brown pigmentation on coxae, lighter shades also on femur and tibiotarsus.
Head. 8+8 eyes, GH smaller than EF ( Fig. 3 View FIGURE 3–8 ). Interocular chaetotaxy with five chaetae (s, t, q, r as mic and p as mes). Holotype antennae length 1.55 mm. Antennal length to head diagonal length ratio 3.23 (holotype). Relation of antennal joints I–IV as 1: 1.8: 1.9: 2.3 (holotype). Ant IV with bilobed apical bulb and bifurcate pin chaeta present ( Fig. 4 View FIGURE 3–8 ). Ant III sensillary organ composed of two sensory rods partially behind a cuticular fold, guarded by three short sensilla ( Fig. 5 View FIGURE 3–8 ). Arrangement of chaetae on labrum 4/554, prelabral chaetae ciliated, posterior, median and anterior labral chaetae smooth. Labrum with four rounded labral papillae with strong, armed spine-like projection ( Fig. 6 View FIGURE 3–8 ). Outer maxillary palp with two smooth chaetae and three smooth sublobal chaetae. Lateral process (sensu Fjellberg 1999) on labial papilla E outreaching apex of papilla ( Fig. 7 View FIGURE 3–8 ). Labium chaetotaxy formed by five smooth “ a ” chaetae and, in the basal row, by ciliated chaetae M,R, E, L 1 and L 2, R reduced ( Fig. 8 View FIGURE 3–8 )
Body. Ratio of Abd IV/III length 5.18–6.36 mm (n=3), holotype 6.36 mm. No differentiated chaetae on tibiotarsus III, with exception of smooth terminal (supraempodial) chaeta opposite to tenent hair. Trochanteral organ with up to 20 smooth spiny chaetae ( Fig. 9 View FIGURE 9–14 ). Unguis and unguiculus of claw III as in Fig. 10 View FIGURE 9–14 . Unguis inner side with sub-equal paired basal teeth at 54 % from inner edge, and with two more unpaired teeth at 74 % and 87 % from inner edge, respectively. Paired lateral teeth intermediate, but at level below the paired internal teeth. Unpaired dorsal tooth basal. A small pretarsal chaeta present on both anterior and posterior surfaces. Unguiculus lanceolate, outer lamella serrated. Tibiotarsal tenent hair clavate, shomewhat shorter than claw. Ratio of supraempodial chaeta / unguiculus around 1. Ventral tube with 9+9 ciliated chaetae on anterior side and 7+7 ciliated chaetae on posterior side ( Fig. 11 View FIGURE 9–14 ); lateral flap with 9 ciliated and 3 smooth chaetae ( Fig. 12 View FIGURE 9–14 ). Manubrial plate with 5–6 chaetae, including two larger inner chaetae and 3–4 chaetae outer two psp ( Fig. 13 View FIGURE 9–14 ). Length of not ringed terminal dens about four times the length of mucro. Mucro with distal tooth equal to anteapical; basal spine present and reaching tip of anteapical tooth ( Fig. 14 View FIGURE 9–14 ).
Macrochaetotaxy ( Figs 3 View FIGURE 3–8 , 15–18 View FIGURE 15–18 ). Simplified Mac formula: 4-1-0-3-1/6(8)-8(9)/2-3(4)/0-2-1/6(8)-6-1-4(5)- 4(5). Head ( Fig. 3 View FIGURE 3–8 ): H1 area with four Mac (An 2, An 3a1, An 3a2, and An 3); H2 area with one Mac (A 5); H3 area without Mac; H4 area with three Mac (S 1, S 3 and S 4i); H5 area with one Mac (Ps 2), Ps 5 present as mes. Mesothorax ( Fig. 15 View FIGURE 15–18 ): area T1 with 6–8 Mac (m 1, m 1e, m 1i, m 2, m 2i and m 2i2 always present, m 2e and m 2pe present or absent); T2 with 8–9 Mac (a 5, m 4, m 4i, m 4i2, m 4p, m 4pi, m 4pi2 and m 5 always present, a 5’ present or absent). Abdomen: Abd II ( Fig. 16 View FIGURE 15–18 ) area A1 with two Mac (a 2 and a 3); area A2 with 3–4 Mac (m 3, m 3e, and m 3ep always present, m 3ei present or absent); Abd III ( Fig. 17 View FIGURE 15–18 ) area A3 without Mac; area A4 with two Mac (a 2 and a 3), and area A5 with one Mac (m 3); Abd IV ( Fig. 18 View FIGURE 15–18 ) area A6 with 6–8 Mac (A 1, B 1, B e2, B e3, D 1 and a Mac of uncertain homology always present; A e2 and another Mac of uncertain homology present or absent); area A7 with six Mac (A 2, A 3, A e3, B 2, C 1 and E 1); area A8 with one Mac (A 4); area A9 with 4–5 Mac (A 5, A e5pp, A i1 and B 5 always present, A e5p present or absent); and area A10 with 4–5 Mac (A 6, A e8, A i2 and B 6 always present, A e7 present or absent); sensillar formula from Th II to Abd V: 2,2/1,2,2,6,3; microsensillar formula from Th II. to Abd III: 1,0/1,0,1.
Ecology and distribution. E. silvicola sp. nov was found in the litter of a mountain beech forest. This new Entomobrya is, therefore, a silvicolous and phytodetriticolous species.
Etymology. The name of the new species refers to the forest habitat of the type locality.
Remarks. E. silvicola sp. nov has a rather unique colour pattern that distinguishes it from other Entomobrya species both in Europe and worldwide. However, with regard to the abdominal macrochaetotaxy, 13 species share the same dorsal macrochaetotaxy formula of Abd II–III ( Table 1 View TABLE 1 ), namely E. amurica Jordana & Baquero, 2021 in Baquero et al. 2021; E. clitellaria Guthrie, 1903 ; E. bicincta Jordana & Baquero, 2021 in Baquero et al. 2021; E. erratica Brown, 1932 ; E. karlstetterae Baquero & Jordana, 2008 ; E. koreana Yosii, 1965 ; E. margaretae Gruia ,
na —information not available.
1967; E. monopunctata Lee & Park, 1984 ; E. nana Lee & Park, 1992 ; E. oleniensis ( Tshelnokov, 1987) ; E. regularis Stach, 1963 ; E. shaanxiensis Jing & Ma, 2023 ; E. tokunagai Yosii, 1942 ; and E. vigintiseta Lee & Park, 1984 , respectively.
Besides the markedly different colour patterns, there are also notable differences in dorsal macrochaetotaxy and other characters between the species mentioned above and the new species ( Table 1 View TABLE 1 ). E. silvicola sp. nov is characterized by a large number of macrochaetae on both Th II area T1 and T2 (6–8 and 8–9, respectively), which differentiates it from nearly all other listed species, with the exception of E. clitellaria and E. margaretae , which have nearly the same number of macrochaetae in the T2 area. The former species has, however, remarkably fewer macrochaetae in almost all Abd IV areas, while the latter species also differs, among other characters, by the higher number of chateae and pseudopore on the manubrial plate. The original description of E. monopunctata and E. tokunagai does not provide information on the chaetotaxy of Th II. However, both species clearly differ from the new species in the number and distribution of dorsal macrochaetae of Abd IV, and in the reduced number (3) of internal teeth on claw III. E. nana has fewer macrochaetae on both Th II area T1 and T2, and only three internal teeth on claw III. The only species that shares the same head macrochaetotaxy is the recently described E. shaanxiensis , but apart from the aforementioned different thorax chaetotaxy, all Abd IV areas (A6–A10) have different numbers of macrochaetae.

| HNHM |
Hungarian Natural History Museum (Termeszettudomanyi Muzeum) |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |