
Aphanius marassantensis, Pfleiderer, S. Johannes, Geiger, Matthias F. & Herder, Fabian, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3887.5.4 |
|
publication LSID |
lsid:zoobank.org:pub:4E7FB25C-6B88-4AF6-8F6C-6DC96B00B55C |
|
DOI |
https://doi.org/10.5281/zenodo.5685455 |
|
persistent identifier |
https://treatment.plazi.org/id/03937F10-F31E-DF43-D1FF-0BF7C99AF8EC |
|
treatment provided by |
Plazi |
|
scientific name |
Aphanius marassantensis |
| status |
sp. nov. |
Aphanius marassantensis View in CoL , sp. nov.
Kızılırmak toothcarp
( Figs. 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 , Tables 1–2)
Holotype. ZFMK 66342 (ex FSJF 3455), male, 32.1 mm SL; Turkey: Ankara prov.: Hirfanlı Reservoir, 39°11'19''N, 33°34'45''E; uncertain collector’s identity (initials B. Y.), 3 May 2008.
Paratypes. ZFMK 66343-66371 (ex FSJF 3455), 29, 14 males, 15 females, 28.3–42.6 mm SL; same data as holotype.
Additional non-type material. ZMH 3480, 86, 19.3–55.2 mm SL; ZMH 3484, 1, 33.2 mm SL; ZMH 3485, 1, 40.1 mm SL; Turkey: Kayseri prov.: Kayseri, Karpuzatan, 38°46'25"N, 35°27'11"E; W. Villwock, 20 May 1959.— ZMH 3481, 33, 17.7–28.2 mm SL; ZMH 26068, 18, 23.0– 34.9 mm SL; Turkey: Kirşehir prov.: Kirşehir: springs in Kirşehir, W. Villwock, 18 May 1959.— ZMH 3482, 20, 17.3–34.5 mm SL; Turkey: Samsun prov.: Balik Gölü, about 12 km east of Bafra, 41°34'45"N, 36°5'00"E; W. Lueken, 19 Aug 1959.
Diagnosis. Aphanius marassantensis was previously included in A. danfordii , but is clearly distinguished from A. danfordii and all other species of Anatolian Aphanius by 11 fixed, diagnostic nucleotide substitutions in the mtDNA COI barcode region ( Table 1). Males larger than 26 mm SL are distinguished from male A. danfordii by having narrower dark-brown lateral bars (antepenultimate bar anterior to caudal-fin base 0.9–1.8 times wider than the anterior white interspace vs. 2.3–4.0). Most males larger than 26 mm SL have an ovoid dorsal profile (vs. an always rhomboid dorsal profile, which is straight up to a vertical line from the pelvic-fin origin in A. danfordii ). Females larger than 30 mm SL are distinguished from A. danfordii females by the absence of vertical rows of darkbrown spots on the caudal and anal fins (vs. 2–3 narrow rows of dark-brown spots, 1–2 rows in the anterior part of anal fin, proximal to the anal-fin base), and 1–3 lateral rows of large dark-brown spots on the flank behind a vertical from the dorsal-fin origin (vs. flank with equally spaced dark-brown spots not organized in rows). Some females of A. danfordii (ZMH 3477) also lack rows of dark-brown spots in the anal and caudal fins. Since the colour pattern on the body of these specimens was faded, we suspect that the spots on the fins too, might have faded away.
nucleotide position relative to A. mento COI
(genbank accession KJ552446 View Materials )
species 0 1 2 2 3 3 3 5 5 6 6
9 6 1 8 5 8 9 1 8 1 2
7 0 7 0 2 5 7 1 9 9 2 A. anatoliae (n=9) A T G T A/T A A A A A T A. asquamatus (n=7) G T A T A A A A G A A A. danfordii (n=6) G T A C A A A A A A T A. fontinalis (n=5) G T A C A A A A A A T A. iconii (n=2) A T A C A A A C C A T A. maeandricus (n=6) A C/T A C A A A A A A T A. marassantensis (n=5) C G C G G C T G C T G A. mento (n=3) G T A T C A A T T A C A. meridionalis (n=8) A T A C A A A A A A T A. saldae (n=2) A T A C A A A A A A T A. sureyanus (n=3) G T A C A A A A A A T A. transgrediens (n=8) A T A C A A A A A A C A. villwocki (n=9) A T A C A A A A A A T
Differential diagnosis. Aphanius marassantensis is distinguished from A. saldae by having a less slender and elongated body shape (BD/SL 28.2–39.6% vs. 20.1–21.3%), complete scale cover (vs. scales absent on back and belly), and a distinctly lower number of scales along the lateral series (25–28 vs. 47–51). It differs from A. anatoliae ( Leidenfrost, 1912) , A. fontinalis , A. iconii , A. maeandricus , A. meridionalis , A. sureyanus ( Neu, 1937) , A. transgrediens ( Ermin, 1946) and A. villwocki ( Figs. 5–6 View FIGURE 5 View FIGURE 6 ) by male and female colour pattern. Male A. marassantensis possess a higher number of dark-brown lateral bars (8–13 vs. 6–7 and 7) than A. meridionalis and A. sureyanus , a lower number of vertical rows of spots on the caudal fin (2–3 vs. 5–6), and a mostly lower number of lateral bars (8–13 vs. 13–19) than A. villwocki ( Hrbek & Wildekamp 2003) . They are further distinguished from A. iconii and A. maeandricus by the coloration of the dorsal fin (black, sometimes with narrow whitish-grey base vs. black with a wide proximal white band and black with large white blotches or a white band), and from A. fontinalis by the coloration of the anal fin (white with 1–3 rows of black spots, sometimes with a black margin, vs. black with a white base). Aphanius marassantensis is distinguished from A. transgrediens by the coloration of the pelvic fins (hyaline vs. fully or partially black), and has narrower dark-brown lateral bars than A. anatoliae larger than 26 mm SL (third-last bar anterior to caudal-fin base 0.9–1.8 times wider than the anterior white interspace vs. 2.1–4.3).
Female A. marassantensis are distinguished from A. sureyanus and A. villwocki by the presence of many darkbrown spots on the flanks (vs. irregular large dark-brown blotches and an almost complete absence of spots), and from A. sureyanus by the coloration of the dorsal fin (hyaline with tiny dark-brown spots on rays and membranes vs. hyaline with few tiny dark-brown spots only on rays). Female A. marassantensis larger than 28 mm SL are distinguished from A. iconii by 1–3 lateral rows of large dark-brown spots on flank behind dorsal-fin base (vs. irregularly set brown blotches and spots), and from A. anatoliae and A. meridionalis by the coloration of the caudal fin (hyaline vs. 1–4 and 1–3 rows of dark-brown spots). They are also distinguished from A. maeandricus and A.
transgrediens by the presence of a single prominent large dark-brown blotch in mid-lateral position on hypural plate (vs. spot in mid-lateral position on hypural plate of comparable size to spots on the flank), and from A. fontinalis by the coloration of the pectoral fins and the anal and caudal fins (hyaline vs. hyaline with black spots).
Description. See Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 for general appearance and Table 2 View TABLE 2 for morphometric data of holotype and 29 paratypes. Dorsal and ventral profiles convex between tip of snout and dorsal- and anal-fin origins, rarely straight; straight or slightly concave along caudal peduncle. Body deepest at about pelvic-fin origin. Greatest body width at opercle or between pectoral-fin base and tip of pectoral fin. Snout rounded, mouth superior, lower jaw almost vertical. Caudal peduncle compressed laterally, 1.0–1.2 (males), 1.1–1.5 (females) times longer than deep. Pectoral fin rounded, pelvic-fin origin 2–5 scales in front of dorsal-fin origin. Anal fin rounded, its posterior tip reaching to a vertical from between the middle and three quarters of caudal peduncle in males and to a vertical of about the middle of the caudal peduncle in females. Dorsal-fin origin 1–5 scales in front of anal-fin origin. Dorsal fin rounded. Caudal fin truncate or slightly rounded. Largest female examined 55.2 mm SL, largest male 37.5 mm SL. Dorsal fin with 9 ½ (7), 10 ½ (15) or 11 ½ (8) branched rays, anal fin with 9 ½ (8), 10 ½ (20) or 11 ½ (2) branched rays. Caudal fin with 7+7 (3), 8+7 (15), 8+8 (6), 9+8 (3), 9+9 (2) or 10+9 (1) branched rays. Pectoral fin with 15 (10), 16 (15) or 17 (5) rays. Trunk and head covered by cycloid scales. Body with 25 (6), 26 (12), 27 (9) or 28 (3) scales along lateral series. Two or three additional scales on caudal-fin base. Total vertebrae number 25–27, of which 10 (1), 11 (8) or 12 (21) are abdominal vertebrae and 14 (12), 15 (16) or 16 (2) complete caudal vertebrae, excluding the hypural complex.
Colouration. See Figures 2–4 View FIGURE 2 View FIGURE 3 View FIGURE 4 . In preservative. Males: Body pale yellow. Upper side of head and back dark brown. In some individuals, scales on back with a dark-brown margin and pale-brown or yellow centre. Flank with 8–13 dark-brown lateral bars. Interspaces and ventral flank pale yellow. Dorsal fin black, in some individuals with a whitish-grey base. Anal fin pale yellow with 1–3 rows of black spots, some individuals with a black anal-fin margin. Pectoral and pelvic fins hyaline. Caudal fin hyaline or cream-yellow with 2–3 narrow vertical rows of black spots. A few black spots or, in some individuals, rows of black spots, at the ventral and dorsal margin of the caudal fin. Vertical rows of black spots on the caudal fin Y-shaped distally in one individual examined. Rarely, two of the dark-brown lateral bars in males are fused dorsally or ventrally, forming a very wide bar with an encapsulated white interspace, resembling the wider bars of A. danfordii . Up to three of the dark-brown lateral bars rarely contain white blotches completely surrounded by the brown colouration.
Females: Body cream- to pale-yellow. Upper side of head and back brown. Scales on back with brown margin and yellowish centre. In some individuals, brown coloration on back reaches down to middle of pectoral-fin origin. Flank with many large dark-brown spots, organized in 2–3 lateral rows on flank behind dorsal-fin base, irregularly set on flank anterior to a vertical of dorsal-fin base. Most individuals with many melanophores on flank. A single prominent large dark-brown blotch in mid-lateral position on hypural plate. Dorsal fin hyaline with small darkbrown spots on rays and membranes. Anal-, pectoral-, pelvic- and caudal fins hyaline.
Colour in life. Males: Body white to silvery white. On dorsal part of the body often with bluish tinge. Pelvic- and anal fins yellow and white proximal to base or completely yellow. Otherwise as for colour in preservative. Females: Body white ventrally, becoming light yellowish to brownish towards the flanks. Otherwise as for colour in preservative.
Distribution and conservation. Aphanius marassantensis is widespread in the Kızılırmak drainage. It and was recorded from the area of Kayseri ( Fig. 7 View FIGURE 7 ) in the upper Kızılırmak drainage down to the delta ( Fig. 1 View FIGURE 1 ). It is known from a wide range of freshwater habitats, ranging from springs and small streams to large dam lakes. The type locality, the Hirfanlı Reservoir, is located on Kızılırmak River (39°11′19″N; 33°34′45″E). Its maximum and average depths are 69 m and 21 m, respectively, and the average area is 219 km 2.
Aphanius danfordii View in CoL has recently been assessed as Critically Endangered in the IUCN Red List ( Freyhof 2014a) due to drainage activities of the Sultan Sazlığı Marshes, and the Kızılırmak population ( A. marassantensis View in CoL ) was already excluded from this assessment. Wildekamp & Valdenburg (1994) mentioned that irrigation canals were constructed between the Kızılırmak drainage and the western part of the Sultan Sazlığı Marshes. Villwock (pers. comm. in Wildekamp & Valdenburg 1994) reported hybridization of the Kızılırmak population ( A. marassantensis View in CoL ) and A. danfordii View in CoL in these canals, but data remained lacking. Due to the much larger range of A. marassantensis View in CoL compared to the highly locally endemic A. danfordii View in CoL , the effect on this species is likely not to be high, but the case deserves monitoring.
There are reports that at some localities in which once founders for aquarium populations of A. marassantensis View in CoL were collected, e.g. “Karpuzatan” in the northern part of Kayseri, suitable habitats were lost due to desiccation (M. Schneider, pers. comm. with first author). Habitat loss also poses a serious threat to other Aphanius View in CoL species, including the widespread A. anatoliae ( Freyhof 2014b) View in CoL .
Etymology. The species epithet is derived from Marassanta, the Hittite name for the Kızılırmak River. An adjective.
Discussion. In the phylogenetic reconstruction presented by Hrbek & Meyer (2003), A. marassantensis shows a sister-group relationship to A. danfordii , a species endemic to the Sultan Sazlığı Marshes at Soysallı in the Develi Depression, which is a small endorheic basin south of Kayseri. Our reconstruction, based on the much shorter mitochondrial COI barcoding fragment, leads to a different topology ( Fig. 8 View FIGURE 8 ), which places A. marassantensis basal to all other Anatolian Aphanius except A. asquamatus ( Sözer, 1942) , which is endemic to Hazar Gölü in eastern Anatolia, well separated from the distribution of the remaining species ( Fig. 1 View FIGURE 1 ). Despite this difference, in both studies A. danfordii and A. marassantensis are each other’s closest relative based on genetic distances derived from the respective molecular marker(s) applied. Interestingly, the K2P distance between the latter two species based on the much longer fragment (11.8% K2P from 2870bps) used by Hrbek & Meyer (2003) is very close to the 11.4% we calculated from the short barcoding region of COI (652 bps). This finding demonstrates once more the suitability of the standard DNA barcode locus COI as a tool suited for the identification of organisms and Aphanius species in particular. The levels of differentiation between the Anatolian species included in this study are very similar to those detected in Mediterranean Aphanius ( Geiger et al. 2014) . The separation of A. marassantensis as a new species is thus well-substantiated also at the molecular level. For further clarification of the deeper phylogenetic relationships, a study using more and also nuclear markers would be necessary. The same applies for the specimens of A. fontinalis from the Salda population ( Fig. 8 View FIGURE 8 ), which might be introgressed by sympatric A. saldae (see also Geiger et al. 2014 Supplemenary Table S2).
While A. villwocki is restricted to the upper Sakarya River, A. marassantensis is the only Aphanius species described to occur in the lower course and delta of a drainage entering the Black Sea ( Fig. 1 View FIGURE 1 ). Hrbek et al. (2002) reported on an Aphanius population from the lower Yeşilırmak River, which is only about 70 km east of the Kızılırmak delta. This population might also belong to A. marassantensis , but no material for comparison was available.
Until the formation of the endorheic basin in the Develi Depression, where A. danfordii occurred in the late Pliocene/early Pleistocene, the area of the contemporary Develi Depression was an open valley, which drained to the İncesu valley, ultimately connected to the Kızılırmak River ( Erol 1999). This fairly recent connection renders the hypothesis that A. danfordii is more closely related to the western and central Anatolian Aphanius species than to A. marassantensis unlikely, and supports the sister-group relationship as in the phylogenetic tree in Hrbek & Meyer (2003).
The killifishes of the genera Aphanius and Valencia from the Western Palaeartic are considered the secondmost threatened group of freshwater fishes in this region ( Freyhof 2014c). The newly described species highlights the importance of conservation measures for distinct populations, which ultimately might turn out to be separate species. While it is essential and most important to attain an improved protection of the habitat and control of invasive species, ex-situ captive breeding can play an important role as a last step to preserve Aphanius species, especially those with a very restricted range, from immediate danger of extinction, e.g. due to rapid habitat degradation. This has already been applied as a conservation action for several species (IUCN 2014). Distributing offspring of a variety of species and populations among interested hobbyists might in the short term help reduce the risk of losing individual strains. The working group AG Eurasier und Nordamerikaner of the German Killifish Association (DKG) successfully follows this principle in the management of its ex-situ breeding programmes and keeps populations from each discrete location separate. Besides their captive breeding efforts, several aquarium hobbyists also support research by providing data and specimens.
The description of this new species in addition to the 11 previously known Anatolian Aphanius species ( Geiger et al., 2014) highlights the character of Anatolia as an integral part of the Irano-Anatolian biodiversity hotspot ( Conservation International, 2005) and will hopefully help stimulate intensified conservation efforts in this special biogeographical region.
Comparative material. Aphanius anatoliae : FSJF 2483, 15, 19.3–36.2 mm SL; Turkey: Aksaray prov.: Spring in Köşk Park within Sultanhanı, 38°14'29"N, 33°32'45"E.— FSJF 2485, 38, 25.1–44.6 mm SL; Turkey: Konya prov.: Meyil Gölü, about 3 km southwest of Esentepe, 37°59'10"N, 33°21'5"E.— FSJF 2527, 9, 21.4–30.4 mm SL; Turkey: Konya prov.: Stream north of Sarıyayla, draining to former Lake Samsam, 39°7'8"N, 32°45'33"E.— FSJF 2610, 4, 21.3–37.8 mm SL; Turkey: Konya prov.: Stream at Gölyazı at road from Eskil to Cihanbeyli, 38°33'9"N, 33°12'3"E. Aphanius danfordii : FSJF 2601, 16, 22.6–41.7 mm SL; Turkey: Kayseri prov.: Spring Soysallı west of Soysallı, west of Develi, 38°23'25"N, 35°21'56"E.— ZMH 3473, 1, 28.9 mm SL; ZMH 3474, 1, 32.1 mm SL; ZMH 3475, 8, 19.4–27.0 mm SL; Turkey: Kayseri prov.: Çayırözü, about 15 km northwest of Develi, 38°25’2’’N, 35°17’22’’E.— ZMH 3476, 21, 19.5–36.4 mm SL; Turkey: Kayseri prov.: Soysallı, 38°20’28’’N, 35°21’47’’E.— ZMH 3477, 33, 19.1–42.9 mm SL; ZMH 3478; 1, 40.1 mm SL; ZMH 3479, 1, 45.9 mm SL; Turkey: Kayseri prov.: Develi, Ilipunar. Aphanius fontinalis : FSJF 3456, 20, 22.1–38.4 mm SL; Turkey: Burdur prov.: Yarışlı Gölü, 37°34’50’’N, 29°56’50"E, from a captive population.— ZFMK 59778, 1, 38.8 mm SL; ZFMK 59793, 1, 32.5 mm SL; ZFMK 59794, 1, 24.8 mm SL; ZFMK 59795, 1, 25.4 mm SL; Turkey: Burdur prov.: Salda Gölü, 37°33'10"N, 29°40'18"E. Aphanius iconii : FSJF 2325, 16, 24.1–32.2 mm SL; Turkey: Isparta prov.: Spring Karaot at shore of Eğirdir Gölü, about 4 km west of Yenice village, 38°8'6"N, 30°54'27"E.— FSJF 2476, 6, 26.0– 36.2 mm SL; Turkey: Isparta prov.: Lower stream Çayköy at Koysazı bridge, southeast of Eğirdir, 37°50'29"N, 30°53'30"E. Aphanius maeandricus : FSJF 1876, 37, 15.8–40.4 mm SL; FSJF 3027, 9, 26.6–39.8 mm SL; Turkey: Denizli prov.: Işıklı spring at Işıklı, 38°19'17"N, 29°51'4"E.— FSJF 2470, 5, 26.2–38.0 mm SL; Turkey: Afyonkarahisar prov.: Spring Düden, 5 km east of Dinar, 38°3'7"N, 30°10' 32"E. Aphanius meridionalis : FSJF 2460, 7, 20.6–36.2 mm SL; Turkey: Antalya prov.: Small field canal south of Kırkpınar, north of Kızılcadağ, 37°8'21"N, 29°55'5"E. Aphanius saldae : ZFMK 59769, 1, 41.6 mm SL; ZFMK 59770, 1, 32.1 mm SL; ZFMK 59771, 1, 30.6 mm SL; Turkey: Burdur prov.: Salda Gölü, 37°33'10"N, 29°40'18"E. Aphanius sureyanus : ZFMK 59775, 1, 25.2 mm SL; ZFMK 59776, 1, 26.5 mm SL; ZFMK 59777, 1, 27.3 mm SL; Turkey: Burdur prov.: Burdur Gölü, 37°43' 55"N, 30°11'49"E. Aphanius transgrediens : ZFMK 59782, 1, 27.9 mm SL; ZFMK 59783, 1, 25.7 mm SL; ZFMK 59784, 1, 29.3 mm SL; ZFMK 59785, 1, 28.1 mm SL; ZFMK 59786, 1, 26.3 mm SL; Turkey: Afyonkarahisar prov.: spring system of Acigöl, 37°49'3"N, 29°55'36"E. Aphanius villwocki : FSJF 2626, 4, 31.0– 55.2 mm SL; Turkey: Afyonkarahisar prov.: spring about 11 km east of Emirdağ, 39°02'53''N, 31°19'38''E.
Material used only in the molecular genetic analysis. With respective NCBI GenBank accession number. Aphanius danfordii : ZFMK 59307, 1, 29.9 mm SL, KJ834535 View Materials ; ZFMK 59308, 1, 28.1 mm SL, KJ834534 View Materials ; ZFMK 59309, 1, 25.3 mm SL, KJ834533 View Materials ; ZFMK 59510, 1, 32.5 mm SL, KJ834531 View Materials ; ZFMK 59511, 1, 28.2 mm SL, KJ834532 View Materials ; Turkey: Kayseri prov.: Sultan Sazlığı Marshes, Soysallı, 38°20'28"N, 35°21'47"E. Aphanius marassantensis : ZFMK 59508, 1, 38.8 mm SL, KJ834528 View Materials ; ZFMK 59509, 1, 30.8 mm SL, KJ834546 View Materials ; Turkey: Kayseri prov.: Kanlı Gölü, Kayseri, 38°41'36"N, 35°19'37"E.— ZFMK 59742, 1, 29.2 mm SL, KJ834545 View Materials ; ZFMK 59743, 1, 31.7 mm SL, KJ834550 View Materials ; ZFMK 59744, 1, 38.0 mm SL, KJ834537 View Materials ; Turkey: Kayseri prov.: Kayseri, Karpuzatan, 38°46'25"N, 35°27'11"E, from a captive population.
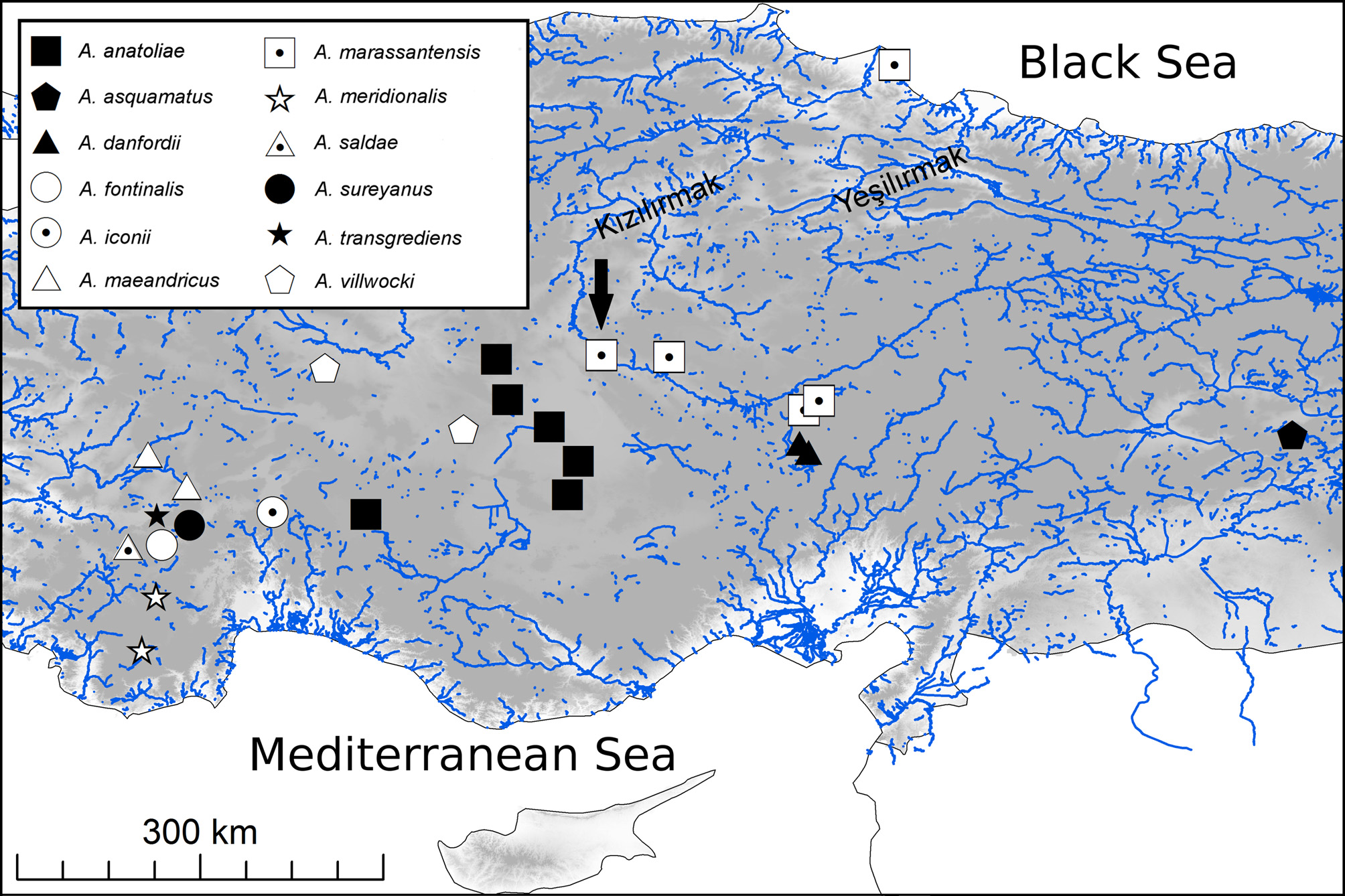






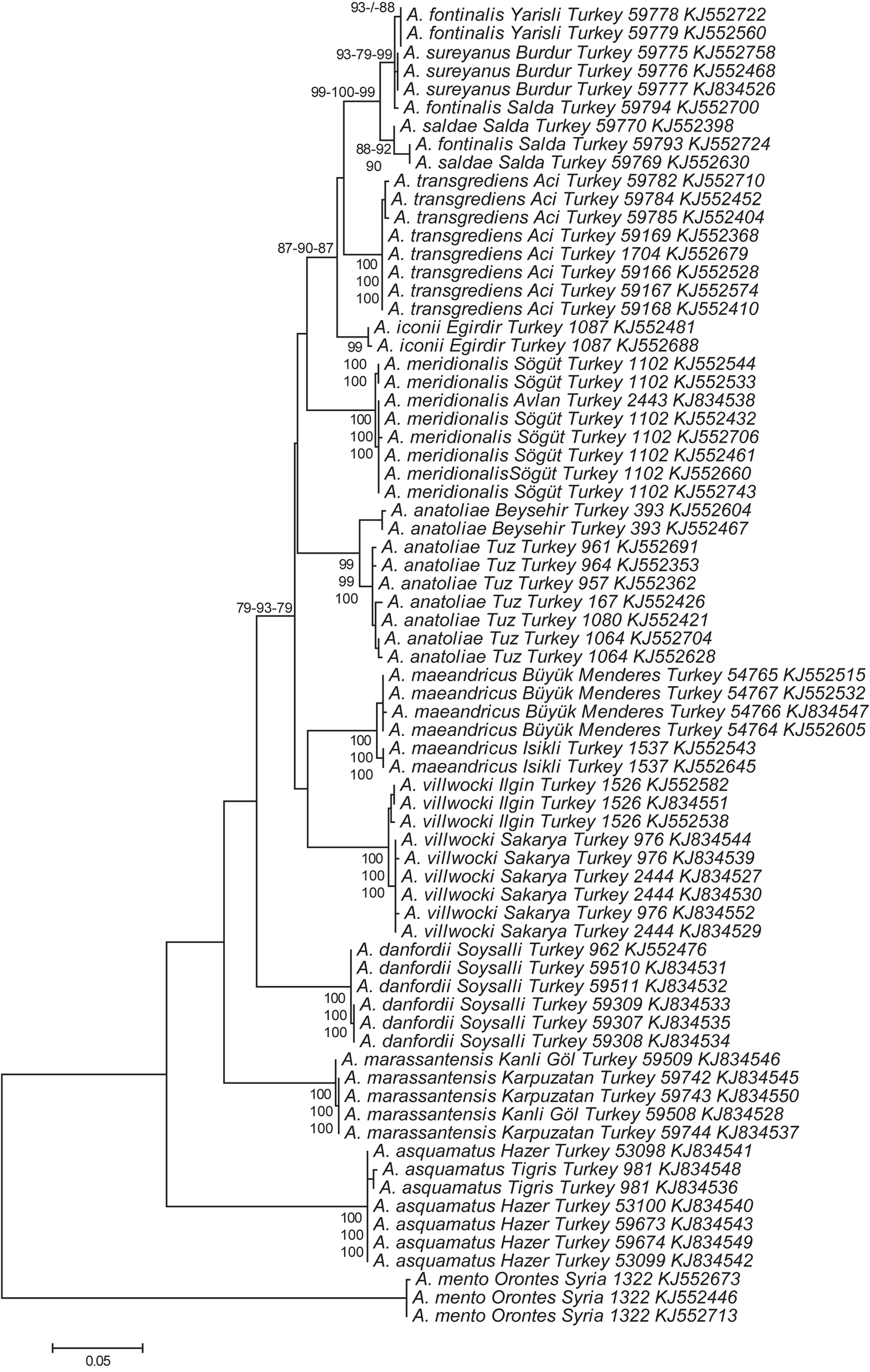
TABLE 2. Morphometric data of Aphanius marassantensis (holotype ZFMK 66342, paratypes ZFMK 66343 – 66371; 29 ex., including holotype).
| males | females | ||||||
|---|---|---|---|---|---|---|---|
| holotype | mean | min | max | SD | mean min | max | SD |
| Standard length (mm) 34.0 | 31.2 | 28.3 | 37.5 | 35.4 31.3 | 42.6 | ||
| In percent of standard length | |||||||
| Head length 30.2 | 30.5 | 27.9 | 32.2 | 1.2 | 29.1 26.5 | 30.8 | 1.1 |
| Body depth at pelvic-fin origin 35.0 | 35.3 | 30.9 | 39.6 | 2.5 | 31.8 28.2 | 35.9 | 2.1 |
| Maximum body width 19.3 | 19.9 | 16.8 | 23.2 | 1.8 | 21.6 19.2 | 24.5 | 1.6 |
| Predorsal length 59.9 | 60.5 | 57.0 | 63.0 | 1.8 | 63.4 61.1 | 65.3 | 1.4 |
| Preanal length 64.4 | 66.3 | 63.6 | 69.9 | 2.2 | 70.8 68.4 | 74.6 | 1.7 |
| Prepelvic length 51.1 | 53.5 | 51.1 | 55.6 | 1.3 | 55.9 52.8 | 59.2 | 2.1 |
| Distance between pectoral and pelvic-fin origins 20.3 | 23.0 | 20.3 | 26.7 | 1.8 | 24.7 20.6 | 26.9 | 1.7 |
| Distance between pelvic and anal-fin origins 13.6 | 14.4 | 12.4 | 17.4 | 1.5 | 16.5 14.9 | 20.0 | 1.3 |
| Depth of caudal peduncle 20.4 | 20.9 | 18.9 | 23.9 | 1.2 | 17.3 16.3 | 18.7 | 0.7 |
| Length of caudal peduncle 23.1 | 22.7 | 21.5 | 23.5 | 0.6 | 22.9 21.3 | 25.0 | 1.1 |
| Dorsal-fin base length 17.0 | 17.4 | 14.6 | 20.1 | 1.5 | 13.6 11.0 | 16.5 | 1.3 |
| Anal-fin base length 15.0 | 13.8 | 10.9 | 16.5 | 1.6 | 10.0 8.9 | 11.3 | 0.7 |
| Pectoral-fin length 17.6 | 17.8 | 16.1 | 20.2 | 1.0 | 15.4 12.6 | 17.1 | 1.0 |
| Pelvic-fin length 12.6 | 12.7 | 12.0 | 14.2 | 0.6 | 10.4 8.1 | 12.1 | 1.0 |
| In percent of head length | |||||||
| Head depth at eye 62 | 64.0 | 61 | 72 | 2.9 | 62.7 59 | 67 | 2.8 |
| Snout length 20 | 18.5 | 16 | 21 | 1.4 | 19.9 17 | 23 | 2.0 |
| Eye diameter 31 | 32.3 | 30 | 35 | 1.6 | 30.1 29 | 33 | 1.4 |
| Postorbital distance 43 | 42.7 | 40 | 45 | 1.5 | 45.3 41 | 48 | 1.9 |
| Maximum head width 61 | 63.4 | 55 | 70 | 4.2 | 70.1 62 | 82 | 5.1 |
| Interorbital width 41 | 40.7 | 38 | 44 | 2.1 | 40.4 38 | 43 | 1.5 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.