
Platypelis tsaratananaensis Guibé, 1974
|
publication ID |
https://doi.org/10.5281/zenodo.215396 |
|
publication LSID |
lsid:zoobank.org:pub:22800BBD-6752-44D0-9335-E760D37C8CA0 |
|
DOI |
https://doi.org/10.5281/zenodo.6176524 |
|
persistent identifier |
https://treatment.plazi.org/id/039387E3-891D-FFB5-FF75-3F84FB2FFA58 |
|
treatment provided by |
Plazi |
|
scientific name |
Platypelis tsaratananaensis Guibé, 1974 |
| status |
|
Platypelis tsaratananaensis Guibé, 1974 View in CoL
Identity. Platypelis tsaratananaensis was described by Guibé (1974) and the holotype specimen MNHN 1993.685 (originally A685) ( Fig. 2 View FIGURE 2 ) was collected in bamboo forest at the Tsaratanana Massif, at 2600 m a.s.l. We reexamined the holotype in 2010. It is an adult specimen of 27.7 mm SVL, sexed by us as a male by its comparatively prominent prepollex (PREPL 1.4 mm, PREPW 0.8 mm) and examination of gonads. The venter of the specimen is cut open, the specimen is in poor state of preservation, and quite weakly fixed. This makes it difficult to take morphometric measurements especially in the region of the head, and many morphological characters therefore cannot be assessed with full certainty. The following account is therefore limited to describing the most relevant characters which we could ascertain and which have not or only incompletely been described by Guibé (1974). Third toe distinctly shorter than fifth toe. Second finger shorter than fourth finger. Tympanum difficult to recognize, probably due to poor state of fixation, its diameter can be estimated about 50% of eye diameter. Coloration almost entirely faded, but above the eyes it is possible to recognize a number of dark lines horizontally slicing a largely light area, which might represent originally yellowish or beige markings with dark marbling. No color pattern visible on the ventral side. Body elongated and hindlimbs relatively short as compared with long body; tibiotarsal articulation reaches between insertion of forelimb and tympanum. For further measurements, see Table 1.
Furthermore, MNHN hosts five paratypes (MNHN 1993.686–690) collected together with the holotype of P. tsaratananaensis . Of these, MNHN 1993.689 (689A) is a juvenile lacking part of the posterior body and the hindlimbs, so measurements are not possible. Specimen MNHN 1993.686 is a subadult of 19.0 mm SVL and has one hindlimb detached but is still relatively complete. The remaining three specimens are adults in moderate state of preservation (but partly in better state than the holotype). MNHN 1993.687 has light markings visible on the flank region, as well as (faintly) lighter color above the eyes than in the surrounding dorsal head surface, and the third toe is shorter than the fifth toe also in this specimen. None of the paratypes has distinct remains of ventral color or any traces indicating a possible pattern of fine dark ventral dotting as mentioned in the original description. However MNHN 1993.687 has some pigment on the throat region which might be a rest of the original color, but no pattern of fine dark dots is visible. MNHN 1993.690 also shows remains of dark stripes of light color in the region above the eyes, similar to the holotype but better visible. We did not check the gonads of the three adult paratypes that we examined; based on the rather comparable size of the prepollex they all might be males, but in any case, none showed distinct characters indicating a female sex, such as oocytes visible through the ventral skin.
Several uncertainties surround the taxonomic status of this species, and we will address these in the following, one by one.
(1) Are any of the Platypelis specimens collected by us on the Tsaratanana massif conspecific with the types of P. tsaratananaensis ?
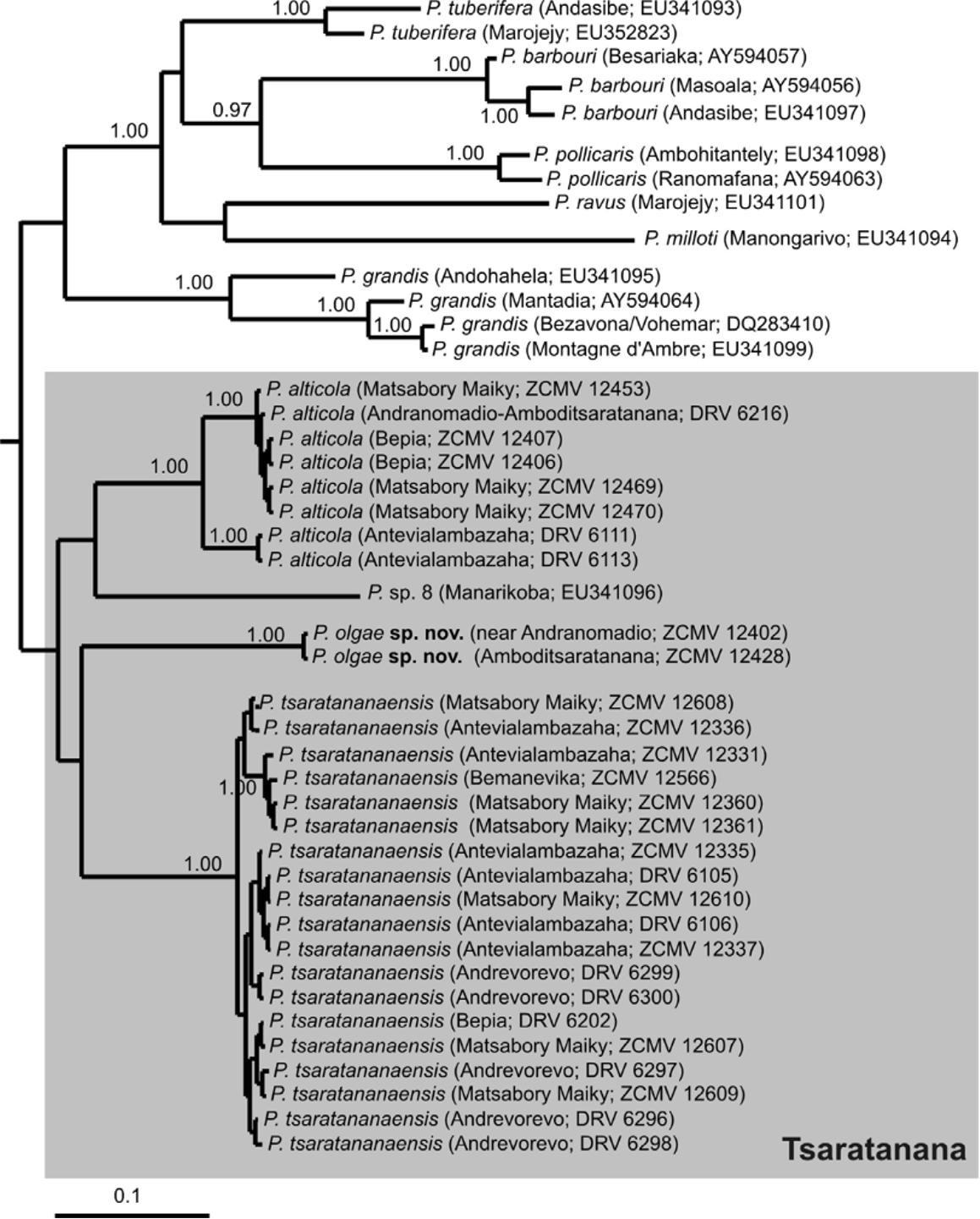
According to our assessment, morphology and color of the P. tsaratananaensis type series, including the namebearing type ( holotype), largely agree with specimens of the most common Platypelis in our collections from the Tsaratanana area. These are similarly sized, with a male SVL of 22–30 mm and a female SVL of 21–33 mm, they have similar relative lengths of toes and fingers ( Fig. 2 View FIGURE 2 ), and a similar elongated body with rather short hindlimbs which differ from most other species of Platypelis . The length of the prepollex of males in life is 2.0 ± 0.2 mm ( 1.5–2.6 mm) and thus slightly larger than in the holotype which however might be due to the poor state of preservation of this specimen ( Fig. 2 View FIGURE 2 ). The remains of light transversal stripes on the head in the holotype and one paratype are indicative of the yellow-beige markings above and between the eyes interrupted by dark lines typical for specimens assigned to this species in life ( Fig. 4 View FIGURE 4 ). Furthermore, the specimens collected by us distinctly differ by genetics and by advertisement calls (see below) from all other nominal species of Platypelis and Cophyla . The only possible difference between our specimens and the type series of P. tsaratananaensis is the purported fine dark dotting of the ventral side of the latter, according to the original description ( Guibé 1974, 1978) which however we could not ascertain in the types while other color patterns still were at least faintly recognizable. The types were collected in bamboo forest ( Guibé 1974,) similar to our specimens, although our collections came from lower elevations ( 1100–2300 m) than the types ( 2600 m). We conclude that the most plausible explanation is to consider our specimens as belonging to Platypelis tsaratananaensis .
(2) Why are our specimens assigned to P. tsaratananaensis not conspecific with Platypelis pollicaris Boulenger, 1888 ?
According to Raxworthy et al. (2008), on the Tsaratanana Massif the species P. pollicaris is very common and occurs from 1650–2450 m a.s.l. This species is morphologically quite similar to P. tsaratananaensis by elongated shape of the body, strong polymorphism of dorsal color, and preference for bamboo habitat, and doubts may arise whether it might be a senior synonym of the latter. Specimens from various localities in eastern Madagascar assigned to P. pollicaris (Glaw & Vences 2007) differ genetically ( Fig. 7 View FIGURE 7. 50 ) and by their single-note advertisement call ( Vences et al. 2006) from the Tsaratanana populations (specimens collected by us emitted a double-note advertisement call—see below), and we are convinced that these eastern populations are correctly assigned to the name P. pollicaris , based on the following rationale: The holotype of P. pollicaris was collected by Rev. R. Baron and described in 1888, without a clear indication of the type locality. As discussed in Ratsoavina et al. (2011), Rev. Baron travelled extensively throughout Madagascar, but apparently reached the northwest, north and northeast of the island only after 1891 ( Dorr 1997). Furthermore, all Malagasy amphibians and reptiles collected by Baron and described by Boulenger with clear type locality information are from type localities in the central eastern portion of Madagascar, far from Tsaratanana. The type of P. pollicaris differs from our Tsaratanana specimens, and agrees with specimens of eastern Madagascar assigned to this name, by two faint but rather constant morphological characters, relative toe length and size of prepollical tubercle in males. First, as stated above, the Tsaratanana specimens (including the holotype of P. tsaratananaensis ), have a third toe distinctly shorter than the fifth toe ( Fig. 2 View FIGURE 2 ). Relative toe length was not mentioned in the original description of P. pollicaris by Boulenger (1888), but Blommers-Schlösser & Blanc (1991) who examined the holotype of P. pollicaris stated that the fifth toe is only slightly longer than the third toe. This agrees with specimens assigned by us to P. pollicaris but less so with the P. tsaratananaensis specimens where the difference between the two toes is strongly marked, not fitting a characterization as faint. Second, P. pollicaris is characterized by a strongly developed prepollical tubercle in males, which probably was even the reason for its scientific name. This tubercle is visible in the holotype drawing of the original description ( Boulenger 1888) and is obvious in specimens from eastern Madagascar that we assign to the species but less distinct in Tsaratanana specimens ( Fig. 2 View FIGURE 2 ). Based on these historical and morphological arguments we conclude that the name P. pollicaris is correctly assigned to specimens from eastern Madagascar (e.g., by Vences et al. 2006; Glaw & Vences 2007), and that based on bioacoustic, molecular and (faint) morphological differences, these populations are not conspecific with our material from Tsaratanana.
(3) Did Raxworthy et al. (2008) observe P. pollicaris at Tsaratanana?
These authors reported P. pollicaris from various sites along the Maromokotro trail, and altogether found 14 specimens from 1650 to 2450 m elevation. We did not re-examine their material and therefore cannot make any definite statement on the identity of this material. However, given that (a) in this elevational range and along the same trail, P. tsaratanananaensis was very common and reproductively active during our own expedition while we never heard the typical call of P. pollicaris in the whole region, (b) Raxworthy et al. (2008) did not assign any other of their specimens to P. tsaratananaensis , (c) P. tsaratananaensis is morphologically very similar to P. pollicaris , we hypothesize that the specimens assigned by Raxworthy et al. (2008) to P. pollicaris may in fact belong to P. tsaratananaensis .
Diagnosis and comparisons. Assigned to the genus Platypelis in the Cophylinae based on enlarged terminal discs on fingers and toes, absence of finger-like prepollex, absence of nuptial pads, occurrence in Madagascar, and molecular phylogenetic relationships. Distinguished from all other cophyline species with enlarged discs on fingers and toes by combination of the following character states: medium body size (adult SVL 22–33 mm), most specimens with colored patches above the eyes, absence of reddish or yellowish ventral color, no sharp border between dorsal and lateral color, toe 3 distinctly shorter than toe 5, vomerine teeth present, males with prepollical tubercle but lacking a finger-like prepollex as typical for Anodonthyla .
Distinguished from all other species of Platypelis and Cophyla by relative toe length (third toe distinctly shorter than fifth toe, vs. only slightly shorter, of equal length, or longer), and by advertisement call (consisting of double-note calls vs. single-note calls). Further distinguished from Platypelis alticola , P. grandis and P. tuberifera by usually smaller body size (adult SVL 22–33 mm but usually below 30 mm, vs. 30–105 mm); from C. occultans , P. ravus , and P. tetra by larger size (SVL 22–33 mm vs. 16–21 mm; from P. barbouri , P. grandis , P. mavomavo , and P. tetra by a completely smooth dorsum (vs. tuberculated), from P. barbouri , P. milloti , P. mavomavo , and P. ravus by always uniform greyish ventral side (vs. reddish or yellow color on venter and/or ventral side of hindlimbs). Furthermore distinguished from all other arboreal microhylids included in our analysis by a substantial genetic differentiation as indicated by mitochondrial DNA (>10% uncorrected pairwise divergence in the 16S fragment studied herein).
Material examined. MNHN 1993.685 ( holotype) and MNHN 1993.686–690 ( paratypes), all collected on Tsaratanana massif (bamboo forest, 2600 m) by R. Paulian in October 1949; ZSM 1638/2010 ( ZCMV 12335, juvenile), ZSM 1639/2010 ( ZCMV 12336), ZSM 1640/2010 ( ZCMV 12337), all collected at Camp 1 (Antevialambazaha); ZSM 1641/2010 (DRV 6105, male), ZSM 1642/2010 (DRV 6107, female), ZSM 1643/2010 (DRV 6129), all collected at Camp 1 (Antevialambazaha) on 10–11 June 2010; ZSM 1644/2010 ( ZCMV 12607), ZSM 1645/2010 ( ZCMV 12610), ZSM 1646/2010 ( ZCMV 12612), all collected at Camp 2 (Matsabory Maiky) on 15–20 June 2010; ZSM 1647/2010 ( ZCMV 12360, male), ZSM 1648/2010 ( ZCMV 12361, female), both collected at Camp 2 (Matsabory Maiky) on 10–11 June 2010; ZSM 1649/2010 ( ZCMV 12391, adult male, call voucher), collected at Camp 2 (Matsabory Maiky) on 12 June 2010; ZSM 1651/2010 (DRV 6202, small juvenile), collected at Camp 3 (Bepia) on 15 June 2010; ZSM 1650/2010 ( ZCMV 12566, female with large oocytes), collected at river crossing near Bemanevika village, 14.48251°S, 48.62723°E, 1109 m a.s.l., on 29 June 2010; ZSM 1652/2010 (DRV 6299), ZSM 1653/2010 (DRV 6300, male, call voucher) collected by F. Ratsoavina at Andrevorevo, 14.3464°S, 49.1028°E, 1717 m a.s.l., on 21 June 2010. Additional specimens were deposited in the UADBA collection and not examined in detail for this study.
Redescription. Because of the poor state of preservation of the holotype, we here provide a redescription based on a newly collected adult male, ZSM 1649/2010 (ZCMV 12391). Specimen in good state of preservation, some muscle tissue removed from right thigh, snout–vent length 26.4 mm. Body very slender; head slightly longer than wide, very slightly wider than body; snout rounded in dorsal and lateral views; nostrils directed dorsolaterally, not protuberant, slightly nearer to tip of snout than to eye; canthus rostralis indistinct; loreal region plain; tympanum distinct, 54% of eye diameter; supratympanic fold very indistinct, virtually absent; tongue ovoid, not bifid or notched; maxillary teeth distinct; vomerine teeth distinctly recognizable as two round groups on a bony ark; choanae rounded. Forelimbs very slender; subarticular tubercles moderately distinct; outer metacarpal tubercle moderately distinct, relatively small and flat; inner metacarpal tubercle large, forming distinct protuberance at base of first finger; hand with only traces of webbing between fingers; fingers distinctly flattened and broad along entire length; relative length of fingers 1<2<4<3, fourth finger distinctly longer than second; finger discs distinctly enlarged, slightly triangular; nuptial pads absent. Hindlimbs slender; tibiotarsal articulation reaching between forelimb and tympanum when hindlimb adpressed along body; tibia length, 40% of SVL; lateral metatarsalia strongly connected; inner and outer metatarsal tubercles small, difficult to recognize; distinct webbing between toe 3 and 4 and less developed between toe 4 and 5; subarticular tubercles on toes moderately well recognizable; toes moderately broad along their entire length; relative length of toes 1<2<3<5<4, toe 5 distinctly longer than toe 3. Dorsal skin smooth, without dorsolateral folds. Ventral skin smooth on throat and chest and moderately granular on belly.
After two years in 70% ethanol, dorsum, thighs and shanks grey with numerous small and well delimited blackish spots; head grey with much less dark spotting; blackish skin above eyes. Forelimbs and hindlimbs without crossbands. Ventrally, throat and limbs uniformly yellowish, belly uniformly greyish.
Variation and distribution. Specimens of P. tsaratananaensis were characterized by a remarkable variability in color and pattern in life ( Fig. 4 View FIGURE 4 ). Of 156 individuals, nine (5.6%) had a yellow to orange vertebral stripe. Most individuals had yellow to beige patches above the eyes which sometimes were large, continuous and contrasting, and sometimes were rather formed as an irregular network of small colored markings. Often a dark line was present between the eyes, and sometimes this black line formed the border of a large light patch on the anterior head. The flanks sometimes had distinct light spotting. The ventral side was always uniform greyish and somewhat translucent. Based on 156 specimens all measured at Matsabory Maiky in the field, males were slightly smaller than females: male SVL was 25.2 ± 1.8 mm ( 22.1–29.8 mm, N= 58) and female SVL was 25.9 ± 2.4 mm (21.0– 32.5 mm, N= 98) (Mann-Whitney U-test, Z=2.281, P = 0.023). All specimens had a smooth dorsal skin without tubercles.
Habitat and natural history. Platypelis tsaratananaensis was almost exclusively found on bamboo, and occurred in high densities in bamboo forest with relatively large plants that we provisionally identified as belonging to the genera Arundinaria and Nastus . Bamboo shrubs were found commonly on the Tsaratanana but at high altitudes the plants were small and the segments probably unsuitable for Platypelis reproduction because of the small diameter of internodes which furthermore did not appear to contain water; P. tsaratananaensis was consequently very rare in these areas. Typically, the species was found in bamboo segments with a small hole, probably made by insects emerging from the bamboo stem after completing their larval development therein, and in some cases opened by the nocturnal lemur Daubentonia madagascariensis ( Rakotoarison et al. 2010) ; especially in living bamboo, these holes were water-filled ( Fig. 9 View FIGURE 9 ).
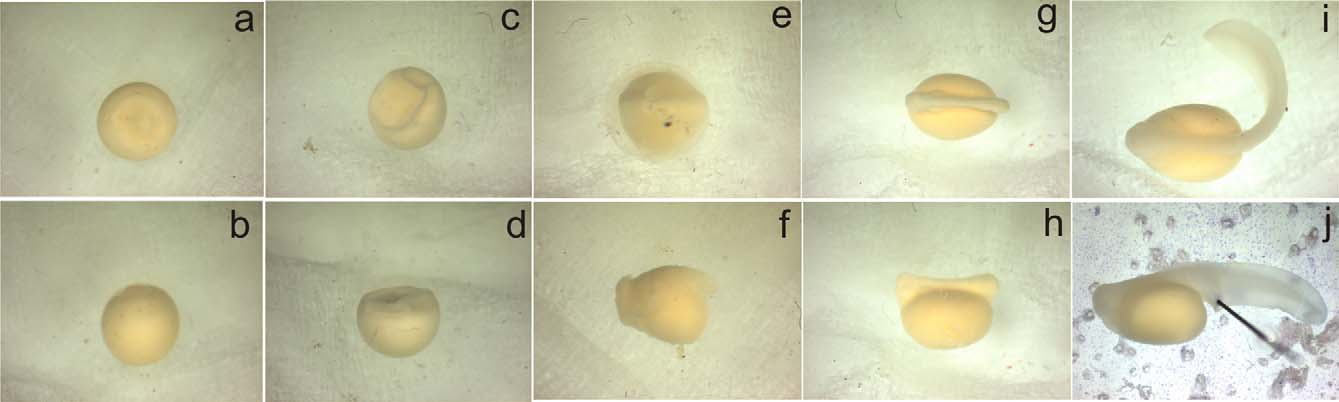
In various cases we observed embryos probably belonging to P. tsaratananaensis ( Fig. 8 View FIGURE 8 ). Sometimes the embryos contained in the same bamboo segment were in distinctly different stages, suggesting that they belonged to separate clutches. The number of embryos found in one segment varied from 1–27 (mean 13 ± 8.1). Adult specimens were observed in bamboo segments of 30.2 ± 7.7 mm ( 30.2–48.9 mm) diameter, located at 151 ± 80 cm ( 30–400 cm) height above the ground. The average number of adult individuals found within one occupied bamboo segment was 2 ± 1 (1–6, N= 87) individuals. Gravid females were larger than non-gravid females (Mann-Whitney test, p = 0.001). We counted 16 ± 5 (1–24, N= 46) oocytes per female. The number of oocytes was correlated with female size (correlation coefficient, r = 0.625, p = 0.001). In those 13 holes where embryos or larvae were found, these were always accompanied with metamorphosed individuals: in the respective segments we found either two males (4 cases), one female and one male (2 cases), one female and 2 males (2 cases), three females and one male (2 cases), one female, one male and one juvenile (1 case), one female (1 case), or three females (1 case). Hence, in most cases the embryos were accompanied by at least one male, and only rarely by only one or several females.
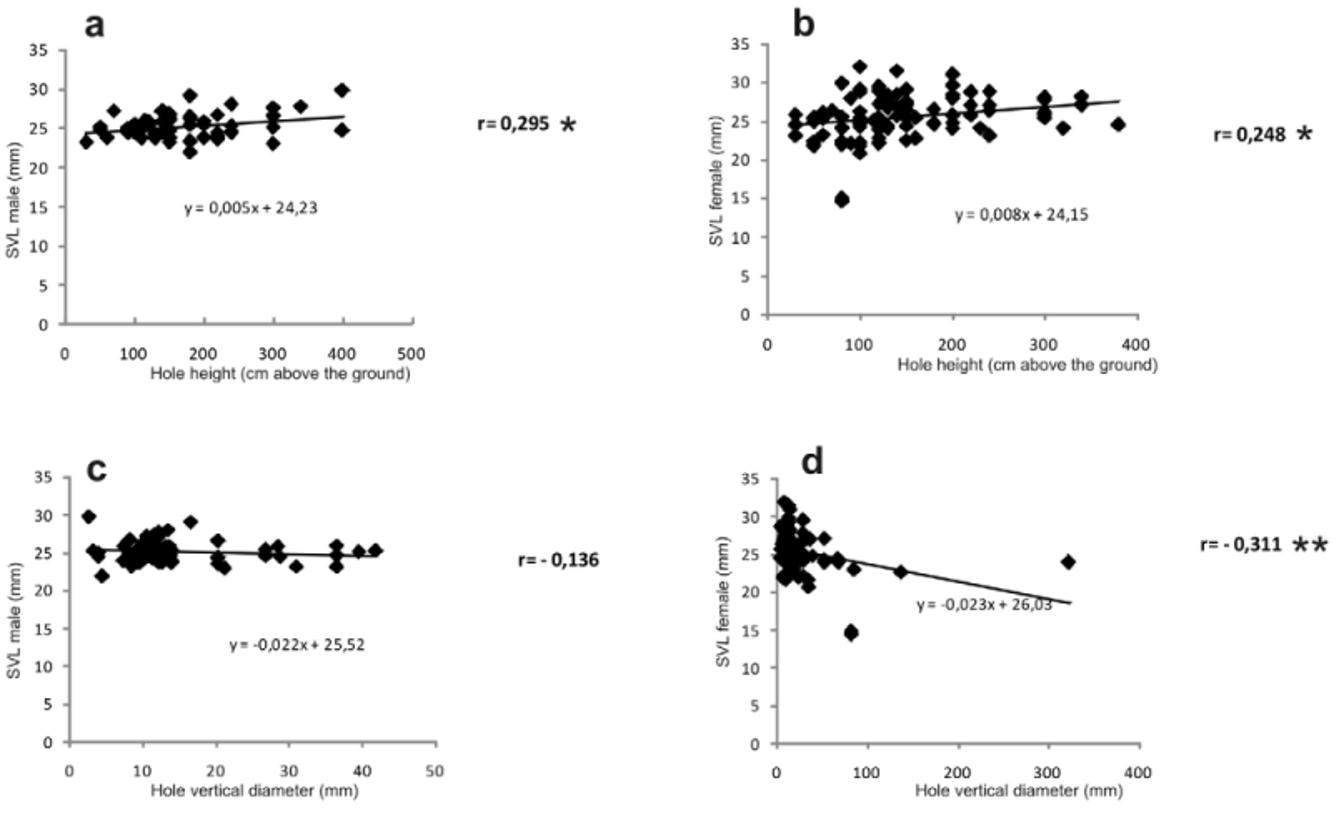
The characteristics of the bamboo segment occupied were related to the size of the frogs found therein ( Fig. 11 View FIGURE 11 ). For this comparison, also some possibly subadult specimens ( 15–21 mm SVL) were included that morphologically could be identified as probably being of male or female sex (based on prepollex size). We found a positive linear correlation between height above the ground and SVL (correlation coefficient, r = 0.295, p = 0.026 for males and r = 0.248, p = 0.012 for females), i.e., larger specimens were found in higher bamboo segments. No correlation was observed between the diameter of the occupied bamboo segment and SVL (correlation coefficient, r = -0.247 with p = 0.64 for males and r = -0.122, p = 0.224 for females). Equally we found no significant correlation between the vertical diameter of the entrance hole to the bamboo segment and SVL of males (correlation coefficient, r = -0.136, p = 0.313), whereas this correlation was highly significant and negative in females (correlation coefficient, r = -0.311, p = 0.002), i.e., paradoxically small females were mostly found in segments with large entry holes. No correlation was observed between the horizontal diameter of the hole and SVL of males and females (correlation coefficient, r = -0.18, p = 0.179 for males and r = - 0.003, p = 0.979 for females).
Many of the encountered specimens of P. tsaratananaensis were infected by mites probably of the genus Endotrombicula (e.g., similar to those visible in Fig. 3 View FIGURE 3 a and 6b for P. alticola ). We recorded 0–54 parasites per individual in males (average 5.3) and 0–14 mites per individual in females (average 3.1). These differences were statistically mildly significant (P = 0.04) in a t-test but not in a non-parametric U-test. The number of mites was not correlated with the size of the specimens (P>0.4).
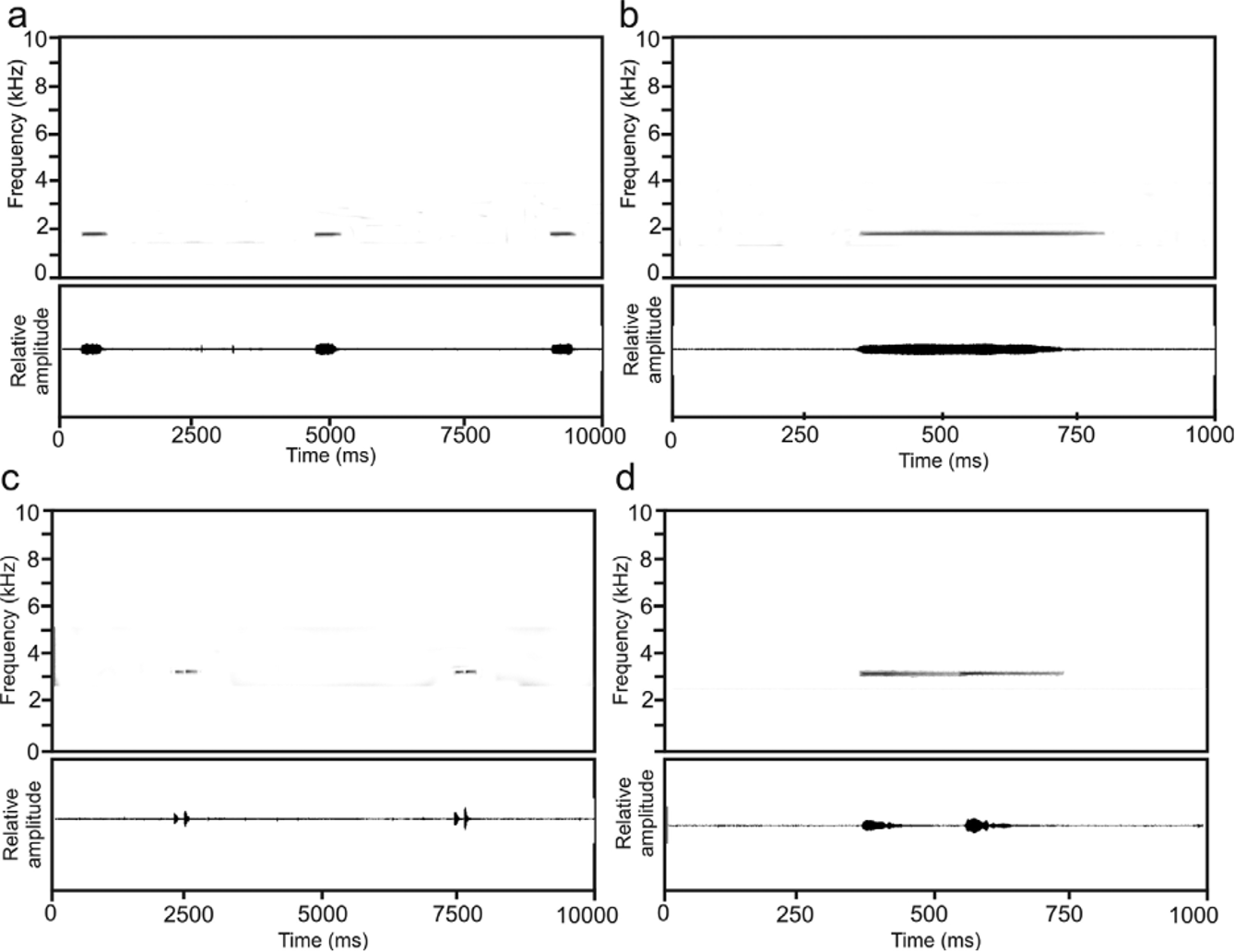
Advertisement call. Calls of Platypelis tsaratananaensis were heard at Matsabory Maiky and Andrevorevo. They are unusual for cophylines in general because each call consists of two distinct notes which however are emitted without a clear silent interval between them ( Fig. 10 View FIGURE 10 ). Calls were recorded from an adult male, ZSM 1649/ 2010 (ZCMV 12391) on 13 June 2010 in the early evening (precise time not recorded, but between 19:00 and 22:00 h) at Matsabory Maiky, at an air temperature of about 13°C. The specimen was not seen calling but clearly located on top of a small palm within a forest with numerous bamboo stands, about 3.5 m above the ground; upon collection it still emitted some faint vocalizations. Spectral call frequency ranged from about 2320–3240 Hz, with a dominant frequency between 3057–3186 Hz (3116±51; N= 8). A weak frequency band could be recognized at 1500 Hz, but it is uncertain whether this represents the fundamental frequency or rather an artefact of the recording. Temporal call parameters are as follows: note length of first note in note pairs 79–100 ms (92±9 ms; N= 4); note length of second note in note pairs 116–145 ms (128±13 ms; N= 4); interval between notes within a note pair 93–110 ms (100±8 ms, N= 4); interval between note pairs 4854–5218 ms (4983±203 ms; N= 3).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |