
Isometopsallops prokopi, Vernoux, Julien, Garrouste, Romain & Nel, Andre, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.195786 |
|
DOI |
https://doi.org/10.5281/zenodo.6204699 |
|
persistent identifier |
https://treatment.plazi.org/id/03938B6C-6211-3046-63F6-E757FEDCFDC0 |
|
treatment provided by |
Plazi |
|
scientific name |
Isometopsallops prokopi |
| status |
sp. nov. |
Isometopsallops prokopi View in CoL sp. nov.
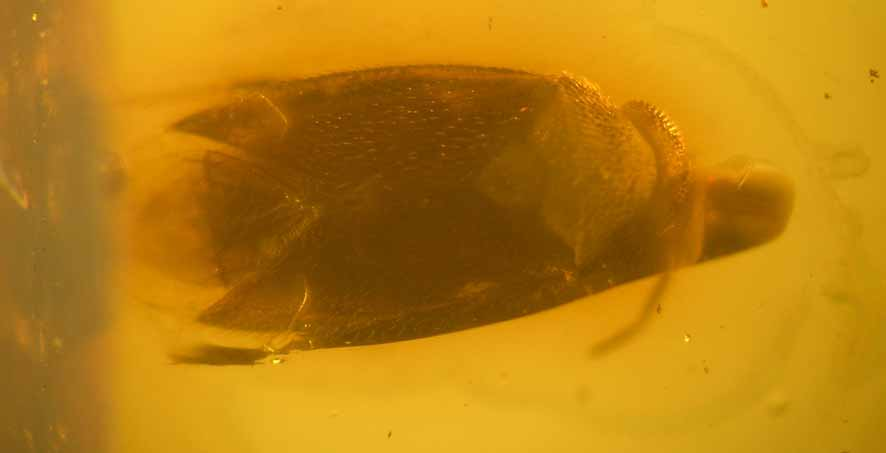
Figs 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4
Material. Holotype MNHN A30851 (PA 8543, male), allotype MNHN A30852 (PA 6524, female), paratypes MNHN A30853 (PA 8584, male), and MNHN A30854 (PA 9493, male), all stored at the Laboratoire de Paléontologie, Muséum national d’Histoire naturelle, Paris, France.
Etymology. Named after our friend and colleague Dr Jakub Prokop (Charles University, Praha).
Diagnosis. tarsi three-segmented; a subapical tooth on each claw; parempodia setiform; no pulvilli; broad femora; seven metafemoral trichobothria and six mesofemoral trichobothria; large eyes semi-globular; antennae placed basally on inner side of eyes; head nearly vertical; no distinct collar; pronotum trapezoidal, ratio length/width pronotum 0.4; membrane of hemelytra crumpled and with two cells, one small and one large; body two times as long as wide, with dorsal surface smooth, with dense hairs; male right genital style (or paramere) slightly longer than left one.
Description. General coloration light brown or brownish; macropterous; dorsal surface smooth, impunctate, with moderately short, erect and sub-erect hairs, densely covering dorsal surface of body and posteriorly directed. Head large, nearly spherical in frontal view, nearly vertical, and nearly twice as wide as long; antenna cylindrical, inserted at inner side of eyes about 1/3 of their lower level; eyes very large, almost semi-globular, covering much of dorsal surface of head, bare, emarginated along inner side of genae; interocular space much narrower than width of eye, hind margins of eyes touching anterior margin of pronotum; mandibular plates not enlarged; labium four-segmented, inserted ventrally on head, and reaching at least a third of abdomen, with segment 1 reaching posterior margin of head; trapezoidal pronotum a little longer than head, with ratio length/width = 0.4, collar absent; no calli; lateral margins and posterior margin not carinate nor emarginated; mesoscutum two times shorter than scutellum; claval commissure present; cuneus and costal fracture clearly distinct; a white spot or light-coloured spot around the costal fracture; hemelytral membrane with a large cell and a small cell which branches to a supplementary vein, membrane distinctly crumpled, hairs absent on membrane; femora of hind legs broad and long, tibia nearly as long as femora, seven metafemoral trichobothria and six mesofemoral trichobothria; tibia with delicate spines distally, three-segmented tarsi more or less of equal size, first tarsal segment well separated from second but suture between third and second segments is weaker than that between first and second (probably presence of a pseudo-trisegmentation sensu Gorczyca [2000]); claws of equal size and almost straight; a subapical tooth on each claw; parempodia setiform; no pulvilli.
Dimension. (taken on PA 8543 and PA 9493 for accessibility): Length of body from apex of hemelytra 2.25 mm (around 2.5 mm for the PA 8584 and PA6524); width 1.025 mm; length of hemelytra 1.75 mm; length of head between 0.25 and 0.375 mm (for different bending of head); width of head 0.625 mm; height of head 0.375 mm; width of eye 0.3 mm; distance between eyes 0.075 mm; antennal segments IV 0.225 mm long, III 0.275 mm long, II 0.4 mm, I 0.115 mm, antennal segment II the longest; length of pronotum 0.35 mm, width 0.875 mm (max.), 0.375 mm (min.); length of mesoscutum 0.15 mm; length of scutellum 0.27 mm; claval commissure 0.525 mm long; costal fracture 0.25 mm long; hind femora 0.87 mm long, (1.05 mm for PA 6524), 0.25 mm wide; hind tibia 0.85 mm long, width mm, tarsal joints I 0.083 mm, II+III 0.180 mm long.
Discussion. After Herczek & Popov (1992) and Popov & Herczek (2006), the mirid subfamily Psallopinae is characterized by the following features: large eyes covering greater part of head, very deeply inserted antennae, absence of ocelli, and presence of subapical tooth on claws. These authors placed the two genera Isometopsallops and Cylapopsallops in this subfamily on the basis of these characters. As they are also shared by Isometopsallops prokopi sp. nov., we consider that it also belongs to this subfamily. One further character that potentially supports an attribution of I. prokopi to the Psallopinae is the presence of seven metafemoral trichobothria, a putative apomorphy also present in the Phylinae , after Schuh (1976: 13).
The Isometopinae , Cylapinae , and Psallopinae share also plesiomorphic characters, i.e., parempodia setiform ( Schuh & Schwartz, 1984; Schuh & Štys, 1991); and absence of pulvilli ( Schuh [1976] observed no pulvilli in Isometopinae , Psallopinae, and Phyllinae ; for Cylapinae , pulvilli are usually present and attached to ventral surface of claw which is an apomorphic character); antennae in inner side of eyes ( Schuh and Schwartz, 1984; Schuh and Štys, 1991).
The Psallopinae is currently considered to be related to the two subfamilies, Cylapinae and Isometopinae ( Gorczyca, 2000; Lin & Yang, 2005). Gorczyca (2000) supported the clade ( Cylapinae + (Psallopinae + Isometopinae )) on the basis of two putative synapomorphies: presence of claws with a subapical tooth, and tarsi two-segmented. The presence of three-segmented tarsi in I. prokopi View in CoL , I. schuhi View in CoL , and Cylapopsallops View in CoL would then be problematic for their attribution to the Psallopinae. Schuh & Schwartz (1984) and Schuh & Štys (1991) considered three-segmented tarsi as plesiomorphic. This character state is present in other mirid subfamilies. However, there are some cylapine genera as Yamatofulvius View in CoL where all the species have apparently functional three-segmented tarsi ( Yasunaga, 2000). Schuh (1976) described also Cylapinae and Isometopinae with tarsi two- or three-segmented (for Isometopinae , see also Herczek [1993]). Thus this character is at least homoplasic. Lastly, Lin & Yang (2005) indicated: "according to Schuh & Slater (1996: 172) tarsi are twosegmented in the Isometopinae and Psallopinae; two- or three-segmented in the Cylapinae ; and threesegmented in the Orthotylinae , Phylinae , Bryocorinae , Deraeocorinae , and Mirinae ." The two-segmented tarsus is judged to be plesiomorphic by these last authors.
The presence of large eyes (also present in the three fossils I. prokopi , I. schuhi , and Cylapopsallops ) would be the main apomorphic characters supporting the clade ( Isometopinae + Psallopinae) ( Schuh & Schwartz, 1984; Schuh & Štys, 1991; Gorczyca, 2000).
Within the clade ( Isometopinae + Psallopinae), the absence of ocelli characterises the Psallopinae and is the unique apomorphy visible on the three fossils I. prokopi , I. schuhi , and Cylapopsallops . For Herczek (1993), it seems obvious that the occurrence of ocelli is plesiomorphic. Schuh & Schwartz (1984) preferred the interpretation of character distributions as a convergent lose of the ocelli in Psallopinae and Cylapinae .
I. prokopi View in CoL differs from the recent Psallops View in CoL in that the right genital style is larger than the left one ( Yasunaga, 1999; Lin, 2004; Lin & Yang, 2005). In recent Cylapinae , the genital styles are asymmetrical in shape and size, one either left or right larger, depending on the species or genus.
Thus nearly all the characters currently used to support the Psallopinae, Cylapinae , and Isometopinae relationships are homoplastic. And so the positions of the three fossils ( I. prokopi View in CoL , I. schuhi View in CoL , and Cylapopsallops View in CoL ) may perhaps be subject to changes in the future.
I. prokopi differs from Cylapopsallops because its body is less elongate (only two times as long as wide, not three times as long as wide); broad femora, not flattened in Cylapopsallops ; collar absent, instead of being distinct in Cylapopsallops .
The habitus of I. prokopi is very similar to that of I. schuhi . They share the same characters mentioned above: tarsi three-segmented (apparently pseudo-trisegmentation); subapical tooth on each claw; trapezoidal pronotum; and two-celled membrane of hemelytra. The ratio length/width of the pronotum = 0.4 in I. prokopi but 0.66 in Isometopsallops schuhi . We lack information on the number of femoral trichobothria in the latter, which renders their comparison for now a little less positive. With the present state of knowledge we prefer to include our fossil in a new species of the same genus Isometopsallops . Unfortunately the male genitalia of I. schuhi are not described.
The presence of a new species belonging to the Baltic amber genus Isometopsallops in the Lowermost Eocene French amber is not surprising because similar fossils occur in other orders (Psocoptera, etc.) ( Nel et al., 2005).
The genus Psallops comprises an undescribed Eocene species from Baltic amber, plus 10 known recent species in the world ( Schuh, 1995; Yasunaga, 1999; Lin, 2004; Popov & Herczek, 2006): P. grandoculus Linnavuori & Alamy, 1982 from Saudi Arabia; P. oculatus Usinger, 1946 from Guam and the Mariana Islands, Saipan; P. ponapensis Carvalho, 1956 and P. yapensis Carvalho, 1956 from the Caroline Islands; P. myiocephalus Yasunaga , P. nakatanii Yasunaga, 1999 , and P. yaeyamanus Yasunaga, 1999 from Japan; P.
formosanus Lin, 2004 and P. l e e a e Lin, 2004 from Taiwan; and P. c h i n e n s i s Lin, 2004 from China. The second recent psallopsine genus Isometocoris is from Panama.
Thus the recent Psallopsinae live under very different climates even if all are warm. One would infer a warm palaeoclimate for the three fossils I. prokopi , I. schuhi , and Cylapopsallops . But as we know nothing about the exact phylogenetic relationships between them and the recent Psallopsinae, and also because the recent representatives of the Isometopinae (putative psallopsine sister group) can be found in very diverse environments and climate, in Europe, North America, Africa, etc., it is in fact not possible to infer anything on the biology of these fossil taxa after their taxonomic position ( Nel, 1997). Nevertheless, recent Psallopinae seem to live on tree trunks and bark ( Yasunaga, 1999). If the corticolous habits may also be inferred for the Eocene Psallopinae, it would not be a surprise to have an overrepresentation of this subfamily in amber inclusions.



| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.