
Azygopus flemingi Nielsen, 1961
|
publication ID |
https://doi.org/ 10.5281/zenodo.215502 |
|
DOI |
https://doi.org/10.5281/zenodo.6174194 |
|
persistent identifier |
https://treatment.plazi.org/id/03945360-4B5D-8D03-FF7F-F96CFB01FAF0 |
|
treatment provided by |
Plazi |
|
scientific name |
Azygopus flemingi Nielsen, 1961 |
| status |
|
Azygopus flemingi Nielsen, 1961 View in CoL
(Figures 1 – 11; Tables 1 – 2) Spotted Flounder
Azygopus pinnifasciatus View in CoL (not of Norman, 1926). — Moreland 1957:34 (off Chatham Islands). Manikiam 1969:120, 124 – 125 (in key; illustration; common name; size; color description; New Zealand waters; depth 190–400 fms; listed, but did not recognize New Zealand subspecies distinct from that in Australian waters). Iwai et al. 1972:29 (brief description, three specimens taken off New Zealand at 270–470 m; mention of subspecies designation by Nielsen 1961). Shuntov 1979:72 (in table; off New Zealand, 400–900 m). Ayling and Cox 1982:313 (to 20 cm TL; color description; color painting; distribution, widespread on continental slope of New Zealand, 300–800 m). Paulin and Stewart 1985:56 ( New Zealand; widespread in deeper coastal waters, 200–600 m). Paul 1986:145 (briefly mentioned in guide to New Zealand fishes; widespread; 300–800 m). Paulin et al. 1989:239, 265 ( New Zealand; in key; illustration). Amaoka 1990:327 (four specimens off New Zealand; size; description with counts, including vertebrae; measurements; color photograph; capture depths 251–551 m; New Zealand and South Australia). Paulin 1996:11–18 ( New Zealand; illustration; non-quota species; distinguished from other New Zealand flatfishes; catch-quota). McClatchie et al. 1997:666 ( New Zealand; listed in Appendix; few catches, low abundance in demersal fish community; 550–558 m). Anderson et al. 1998: unpaginated map ( New Zealand; geographic and bathymetric distributions; relatively rare, collected in only 20 of 19232 stations). Jacob et al. 1998:2127, 2131 (member of demersal fish fauna of southern New Zealand; captured in relatively low abundance). Paul 2000:145 (after Paul 1986). Paulin et al. 2001:239, 265 (re-issue of Paulin et al. 1989).
Azygopus pinnifasciatus flemingi View in CoL — Nielsen, 1961:221, Figure 3 View FIGURE 3 ( New Zealand; original description; three specimens; illustration; type locality Tasman Sea off Greymouth, South Island; 610 m). Whitley 1968:47 (listed as Bothidae View in CoL ; holotype and paratype in Zoological Museum, Copenhagen; second paratype in Australian Museum). Nielsen 1974:84 (holotype and one paratype listed in type catalogue). Sakamoto 1984:210 (listed in checklist of species of Pleuronectidae View in CoL ). Evseenko 2004:19 (in annotated checklist; type specimen and type locality; junior synonym of A. pinnifasciatus Norman View in CoL ). Eschmeyer & Fricke 2012 (junior synonym of A. pinnifasciatus Norman View in CoL ).
Diagnosis. Azygopus flemingi is readily distinguished from the closely-related A. pinnifasciatus by its conspicuous ocular-side color pattern consisting of numerous, small, rusty-brown, nearly spherical spots on a yellowish-white to light-brown background (vs. darker brown ocular-side background color with several darker, irregular, large blotches), in having fewer ocular-side pelvic-fin rays (6–10, usually 9 or fewer vs. 9–12, usually 10 or more in A. pinnifasciatus ); more total vertebrae (46–49, modally 47–48, vs. 44–46, modally 45–46); fewer total gillrakers (11–16, usually 13–15 vs. 13–20, usually 15 or more) and fewer gillrakers on the lower limb of the anteriormost gill arch (8–11, usually 9 or 10 vs. 10–14, usually 10–12 in A. pinnifasciatus ); in having the ventral margin of the lower eye juxtaposed against the middle and posterior regions of the dorsal borders of the ocular-side maxilla and premaxilla (i.e., no space between eye and jaw; compare Figures 3 View FIGURE 3 A and 3B) and in having the posterior border of the jaws only slightly angled away from the orbital rim (vs. ventral margin of lower eye separated from mid and posterior regions of dorsal border of ocular-side maxilla and premaxilla by space containing wedge of 1–3 scales between jaw and orbit, and with posterior border of jaws more steeply-angled away from orbital rim), by its longer (vs. shorter) blind-side pectoral fin, as well as by its uniformly whitish blind-side pigmentation without small, pepperdot-size, reddish-brown spots (vs. blind side of A. pinnifasciatus with numerous, small, reddish-brown, pepperdot-size spots, especially dense on body regions overlying proximal pterygiophores of dorsal and anal fins; compare Figures 4 View FIGURE 4 A and 4B), and by the pigment pattern on dorsal and anal fins (1-2 pigmented rays alternating with 1-2 lighter, but still pigmented, fin rays vs. 3-5 pigmented rays alternating with about 10 lighter or unpigmented rays in A. pinnifasciatus ). Mid-body scales on the blind side differ in these two species (compare Figures 4 View FIGURE 4 A and 4B). In A. flemingi , these scales are relatively close-set compared with those of A. pinnifasciatus , and these scales have two alternating rows of marginal ctenii covering nearly the entire posterior scale margin (vs. ctenii more restricted to mid-region of posterior margin of scales in A. pinnifasciatus ). Proximal and distal portions of exposed areas of these blind-side scales in A. flemingi are asymmetrical in size, with the proximal region usually consisting of 30% or less of the exposed area, and distal regions of exposed areas on these scales (70–85% of exposed area) almost entirely covered by a crescent-shaped area consisting of the medial and basal zones (vs. proximal areas on blind-side scales usually much larger than distal regions, and the distal region with wedge-shaped medial and basal zone in A. pinnifasciatus ). The anterior margin of the calcified plate on blind-side scales of A. flemingi lies nearly in a straight line along its dorsal-ventral axis, whereas the anterior margin of the calcified plate on these scales in A. pinnifasciatus is noticeably angled with an anterior projection medially.
Description. Meristic features summarized in Table 1, with values for the holotype indicated. Dorsal-fin rays 95–110. Anal-fin rays 78–93. Caudal-fin rays 17–19, usually 18 (49/ 53 specimens). Ocular-side pectoral-fin rays 6–12, usually 10–12 (44/ 49 specimens). Blind-side pectoral-fin rays 5–10, usually 6–9 (45/ 49 specimens). Ocularside pelvic-fin rays 6–10, usually 9 or less (51/ 54 specimens). Blind-side pelvic-fin rays 4–6, usually 5 (42/ 53 specimens). Abdominal vertebrae 9–11, usually 10–11 (typically 3 vertebrae without hemapophyses and 7–8 with hemapophyses). Total vertebrae 46–49, modally 47–48 (38/ 55 specimens). Supracranial dorsal-fin proximal pterygiophores usually 7–9 (50/ 52 specimens), rarely 6 (2/52). One dorsal-fin proximal pterygiophore (52/ 52 specimens) associated with first neural spine. Dorsal-fin proximal pterygiophores inserted into first interneural space 2–7, usually 3–5 (50/52 individuals), rarely 2 or 7 (1 individual each with these counts). Dorsal-fin proximal pterygiophores inserted into second interneural space 2–4, usually 2–3 (51/52 individuals). Number of interneural spaces where 3 proximal dorsal-fin pterygiophores insert 3–13; interhaemal spaces where 3 proximal anal-fin pterygiophores insert 2–10. Number of anal-fin proximal pterygiophores inserted anterior to first haemal spine 9–15, usually 12–13 (37/ 52 specimens). Lateral-line scales 95–113. Gillrakers on upper limb of anterior gill arch 3–6, modally 4–5 (37/ 50 specimens). Gillrakers on lower limb of anterior gill arch 8–11, modally 9–10 (47/ 52 specimens). Total number of gillrakers on anterior gill arch 11–16, modally 13–15 (41/ 50 specimens).
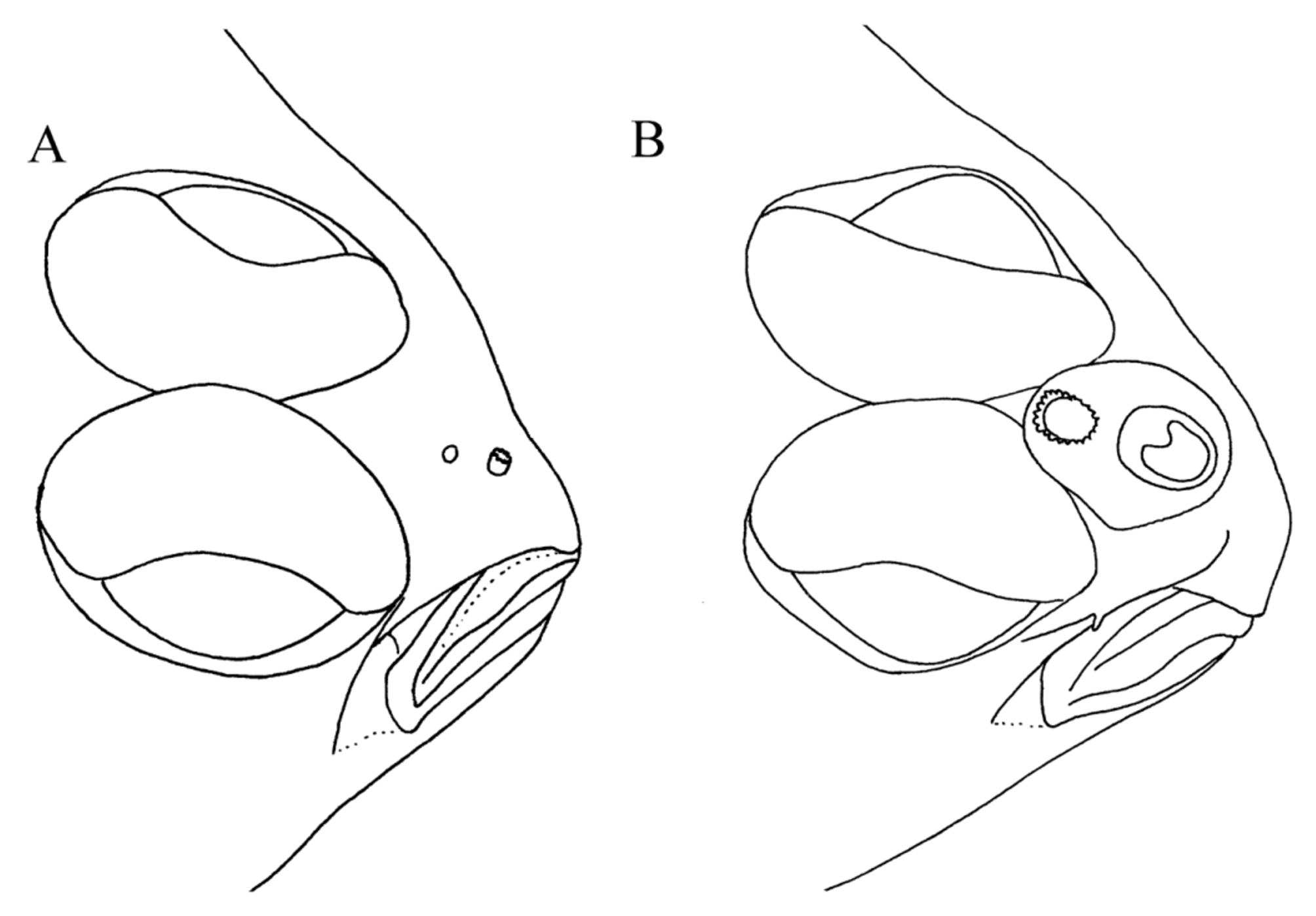
Morphometric features summarized in Table 2. Body ovate, laterally compressed (Figure 1); greatest body depth (34.9–48.7% SL) anterior to body midpoint (usually at vertical between anal-fin rays 10–15), with noticeable taper posterior to this point and gradual taper anterior to this point. Caudal peduncle short. Head small (20.8–28.4% SL), averaging 4.2 times in SL; with slight notch on dorsal profile just anterior to anterior margin of upper eye. Snout short, much shorter than eye diameter (average 2.1 times in ED), slightly to bluntly pointed. Ocular side of snout with scales reaching nearly to anterior margin and with small, triangular-shaped, wedge of scales on snout region posterior to nostrils, the apex of this triangle extending posteriorly into interorbital space. Blind side of snout, except for tip, covered with small ctenoid scales. Two nostrils on both sides of head; ocular-side nostrils more conspicuous than blind-side counterparts. Females with small, inconspicuous nasal sac, and with ocular-side posterior nostril about same size as, or slightly smaller than, anterior nostril ( Figure 2 View FIGURE 2 A); males with conspicuous nasal sac and with large posterior nostril (compared with that of females of comparable size) surrounded by more conspicuous fleshy tube about equal to length of opening ( Figure 2 View FIGURE 2 B). Ocular-side nostrils located closer to horizontal through dorsal margin of lower eye than to horizontal through ventral margin of upper eye. In both sexes, ocular-side anterior nostril a short, round tube with opening located dorsal to anterior third to midpoint of upper jaw and surrounded by short, fleshy tubular nostril with an extended wide flap posteriorly; length of nostril about twice that of diameter of nasal opening. Posterior ocular-side nostril in both sexes a rounded to sphericalshaped opening, with or without slightly raised circular rim located slightly posterior to vertical through midpoint of upper jaw. Blind-side nostrils in both sexes smaller, more translucent, and less conspicuous than ocular-side nostrils; located on snout antero-ventral to dorsal-fin origin. Anterior blind-side nostril a small opening with low, fleshy rim; posterior blind-side nostril with opening oriented dorso-ventrally and surrounded by fleshy rim posteriorly. Mature males with prominent, enlarged nasal capsule on blind side of snout. Blind-side nostrils much larger than those of females. In males, opening of blind-side posterior nostril as wide as length of anterior nostril; females with opening of blind-side posterior nostril much smaller, about equal to only one-fifth length of anterior nostril. Males with base of first dorsal-fin ray usually posterior to, or at, vertical through posterior margin of blindside nasal capsule; females with base of first dorsal-fin ray usually at, or slightly anterior to, vertical through posterior nostril. Eyes large (31.7–53.3% HL), conspicuous, elliptical (ca. 1.8 times longer than wide); upper eye slightly in advance of lower, or eyes symmetrical in position; upper eye very close to dorsal margin of head with only 2–4 small scales between dorsal margin of eye and dorsal margin of head at base of dorsal fin; eyes nearly contiguous throughout anterior one-third to one-half of their lengths; upper aspect of eyes, except for pupils, covered with small, ctenoid scales; eyes without pupillary operculum; outer margins of eyes surrounded by fleshy orbital rim. Interorbital space present between posterior one-half to two-thirds of eyes, narrow, usually without scales or occasionally with single row of ctenoid scales in posterior extent of interorbital space. Mouth small (upper jaw length 22.0–33.3% HL), smaller than eye diameter; jaws angled slightly ventro-posteriorly; posterior end of jaws slightly curved ventrally; lower jaw with small, bony protuberance on symphysis; posterior margin of maxilla reaching point between verticals through anterior margin of iris and anterior margin of lower eye. Dorsal margin of maxilla from about its midpoint to posterior margin nearly touching ventral margin of orbit ( Figure 3 View FIGURE 3 A); no scales between dorsal margin of posterior half of maxilla and ventral rim of orbit ( Figure 3 View FIGURE 3 A); scales, if present between. size sample = N; holotype the of those are asterisk with Values. pinnifasciatus . A and flemingi Azygopus of features meristic for counts of Distribution. 1 TABLE orbit and jaw, only in lachrymal region at, or just beyond, posterior end of maxilla. Jaws with small, acute teeth. Ocular-side premaxilla and dentary with few, small teeth only on anteriormost regions. Entire blind-side jaws with stronger teeth than those on ocular-side jaws. Nearly entire blind-side premaxilla with four rows of slightly recurved teeth, narrowing to two rows at anterior and posterior ends of bone; teeth on anterior end of blind-side premaxilla longer than those at posterior end. Blind-side dentary with large, strong, recurved, teeth over entire jaw; teeth in 2–3 rows medially, narrowing to 1 row anteriorly and posteriorly. Ocular-side head regions dorsal, ventral, and posterior to eyes, with strongly ctenoid scales; ventral side of jaws with single to double row of ctenoid scales extending nearly to anterior tip. Ocular-side preopercle covered by skin in many specimens and not clearly visible; blind-side preopercle with ventral region free and not covered by thick skin. Postorbital head length short (9.6–13.4% SL). Blind side of head with strongly ctenoid scales from opercular margin nearly to snout tip. Gill covers strongly joined to each other, but free from isthmus. Gillrakers short, conical, sharply pointed at tip. Gillrakers on upper limb of blind-side anterior arch nearly equal in length to those on lower limb; those on lower branch of blind-side anterior arch short, wider at their bases; some gillrakers with pointed tips, others with rounded or squarish tips.
Lateral line developed on both sides of body; forming slight curve anteriorly, and continued onto caudal fin posteriorly. Anus situated just off midline on left side of body. Pelvic fins and membranes surround anterior half of anus. Genital pore a small, thick, white tube located on right side of body about 1–2 scales dorsal to, and slightly posterior to, ventralmost ocular-side pelvic-fin ray (genital pore not obvious in some specimens).
Scales relatively small, ctenoid on both sides of head and body. Scales in mid-body region on blind side of body relatively close-set ( Figure 4 View FIGURE 4 A), with two alternating rows of strong ctenii along nearly entire posterior margin of scale. Proximal and distal portions of exposed area of scales asymmetrical in size, with proximal region usually consisting of 30% or less of exposed area. Distal regions of exposed area on these scales nearly entirely covered (70–85% of exposed area) by crescent-shaped area consisting of the medial zone (formed by 1–2 rows of truncated ctenii) and basal zone (consisting of calcified plate formed by fused bases of truncated ctenii and subctenii). Anterior border of calcified plate forming nearly a straight line along its dorsal-ventral axis.
Dorsal-fin origin slightly on blind side of head anterior to vertical through anterior margin of upper eye and dorso-posterior and slightly posterior to blind-side nostril; first few dorsal-fin rays also based on blind side of head. First dorsal-fin ray elongate, separate from second dorsal-fin ray, and noticeably thicker than next few rays. [Figure of holotype appearing in Nielsen (1961) indicating first dorsal-fin ray connected to second ray via membrane in error.] Second dorsal-fin ray more slender and shorter than successive dorsal-fin rays. Anteriormost 5–6 dorsal-fin rays shorter than remainder of dorsal-fin rays, but increasing successively in length posteriorly. Beginning with second dorsal-fin ray, anteriormost 2–10 rays with membrane only on basal half; more posterior rays with membrane nearly to distal tips and with short, distal tips free from membrane. Dorsal and anal fins free from caudal fin. Anal and dorsal fins with sheath of scales covering basal portions of fins, but without scales on fin rays. Pectoral fins developed on both sides of body; asymmetrical; ocular-side pectoral fin nearly twice as long as blindside counterpart. Both pectoral fins with several dorsalmost interior rays longer than marginal rays and other interior rays; no rays elongate. Ocular-side pectoral fin with several, small scales on its base. Pelvic fins developed on both sides of body; asymmetrical, with ocular-side fin nearly twice as long as blind-side counterpart. Pelvic fins with membranous connection to body posteriorly, but free from each other and free from anal fin. Distal tips of pelvic-fin rays free from membranes. Ocular-side pelvic fin with middle rays longer than other rays; origin of ocular-side pelvic fin at tip of isthmus and with its anterior rays situated nearly on ventral midline with more posterior rays inserted gradually more dorsally on right side above midline and positioned noticeably more dorsal than anterior rays. Blind-side pelvic fin short-based, usually with 5 rays; its origin about opposite base of seventh ray of ocular-side pelvic fin. Blind-side pelvic fin with anterior rays closer to midline and with posteriormost rays positioned more dorsally away from midline, but not as noticeably as those of ocular-side pelvic fin. Caudal fin relatively large, approximately 4.5 times in SL, rectangular with central rays slightly longer than marginal rays. Central caudal-fin rays branched, marginal rays unbranched. Both sides of caudal-fin rays with ctenoid scales nearly covering their entire lengths.
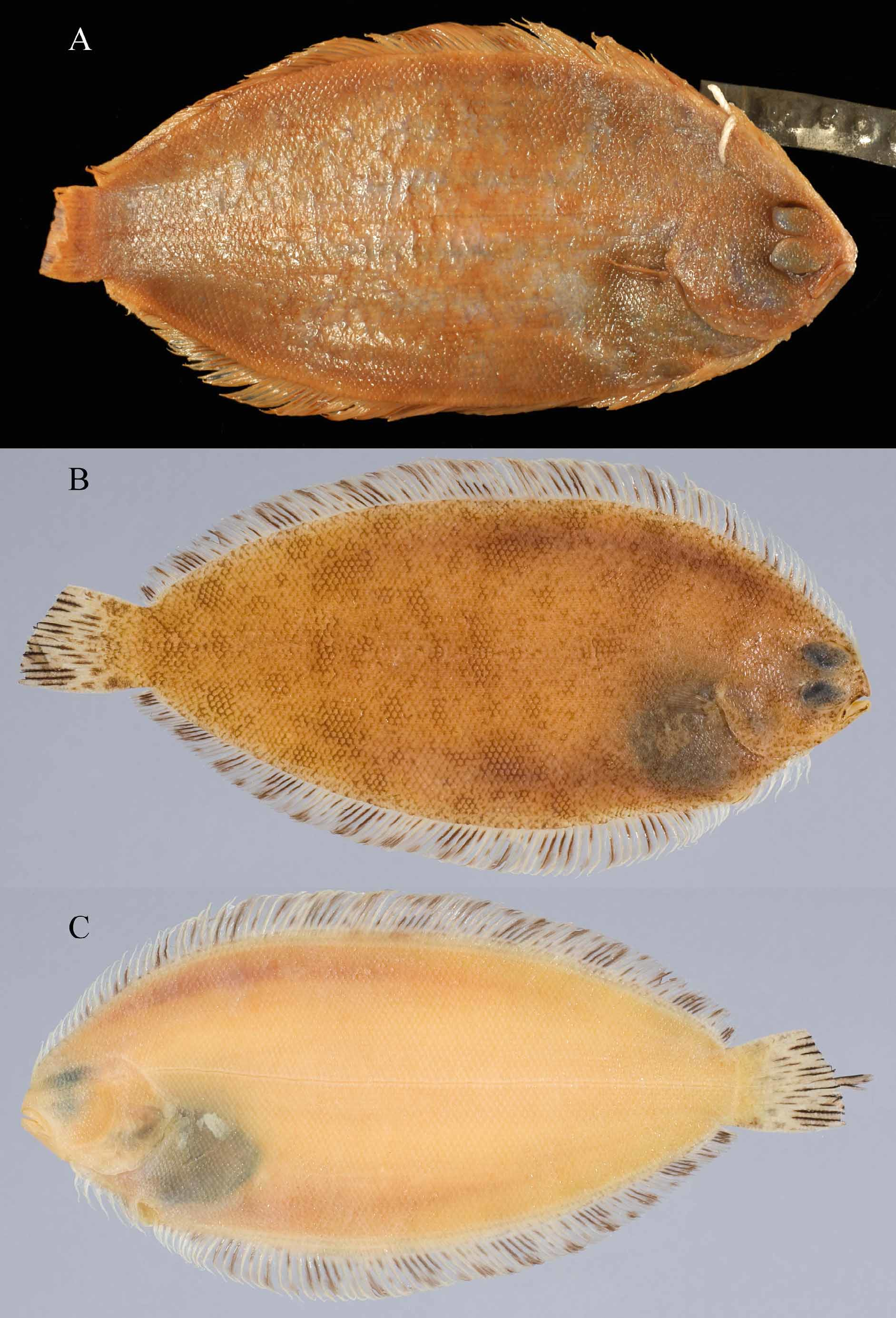
Color in Preserved Large Juveniles and Adults ( Figures 5 View FIGURE 5 A–C). Background coloration of ocular side of body yellowish-white to light-brown or reddish-brown, with numerous, conspicuous, irregular to spherical, darker rusty-brown spots covering nearly entire head and body, and with several larger (some about equal to 0.5 eye diameter), spherical to sub-spherical, (sometimes diamond-shaped), faint blotches on body (often difficult to see amongst rusty-brown spots in larger specimens); and a black peritoneum, visible through abdominal wall on both sides of body, usually more conspicuous on blind side; some specimens also with intense black blotch on anterodorsal body cavity beneath ocular-side pectoral fin. Some specimens with impression of faint, dusky, evenlyspaced, circular spots along body at bases of dorsal and anal fins (see Figure 6 View FIGURE 6 A). Individual ocular-side scales also with numerous, small, reddish-brown melanophores. Scale pockets not outlined in darker pigment than that occurring on scales; some specimens with darker posterior margins on scales corresponding to underlying areas in dermis with rusty-brown spots. Larger juveniles and adults with rusty-brown spots from anterior margin of snout, on upper aspects of eyes, over entire posterior head and body, and extending posteriorly onto basal third of caudal fin. Rusty-brown spots on head and body of uniform size and intensity, not aligned with individual scales, but often offset from center of posterior scale margin, and underlying portions of more than one scale. Anterior snout also with dusky, blackish-brown pigment, several faint blotches, and numerous small, pepperdot melanophores. Ocularside upper and lower jaws black, except tip of dentary white. Ventral margin of lower jaw with numerous melanophores. Ocular-side nostrils translucent. Inner lining of mouth white. Eyeballs grayish-blue to bluish-black with numerous, small, brownish spots on upper aspects. Inner lining of ocular-side opercle yellowish-white, without black pigment. Inner lining of blind-side opercle unpigmented. Gill filaments white.
Ocular side of dorsal fin with anteriormost 4–5 fin rays unpigmented; more posterior fin rays and all anal-fin rays pigmented. Dorsal and anal fins of large juveniles and most adults with pattern of darkly-streaked fin rays alternating with 1–2 lighter (but still pigmented) fin rays. Streaked fin rays in each fin with dense patches of brown melanophores on their basal thirds and with fewer melanophores extending along nearly entire length of fin ray. Darkly-pigmented fin rays sometimes completely streaked over their entire length or with dark pigment only on basal half or distal halves of fin ray.
Ocular-side pectoral-fin rays with brown melanophores along their entire lengths. Membrane between fin rays mostly hyaline. Distal one-third of fin rays darker than basal two-thirds. Fleshy base of pectoral fin with greater concentration of melanophores than that on fin rays. Ocular-side pelvic fin sometimes with all fin rays pigmented, otherwise this fin with anteriormost two fin rays white and with successive fin rays alternating between light and heavy pigmentation. Pigmented pelvic-fin rays anteriorly with small number of brown spots basally, more posterior fin rays with darker pigment over their entire lengths. Blind side of ocular-side pelvic fin with numerous, small melanophores on basal half of fin and with small melanophores extending along lengths of some fin rays, especially those located more posteriorly in fin.
Both sexes with two large, conspicuous, black, circular to elongate blotches on dorsal and ventral margins of posterior region of caudal fin; spots cover both fin rays and membrane. Spots more darkly pigmented on ocular side of caudal fin, but also visible on blind side of fin. Some specimens (e.g., NMNZ P.5806, N=7) with black caudal spots ringed with white pigment or with white border on external margins of dorsalmost and ventralmost fin rays. Caudal-fin spots of males usually darker and slightly larger than those of females, and males, in general, with more heavily pigmented caudal fin than that of females, but neither sex with dark black streaking along lengths of caudal-fin rays. Basal and medial sections of caudal fin also with 1–2 rows of about 3–4, fainter, nearly spherical spots about equal in size to 0.5 ED and each covering about 3–4 fin rays. Each row and each spot separated from next by 1–2 scales. Another irregular row of heavier pigmented blotches mainly on the fin rays about at the mid-fin region. Numerous smaller melanophores present also along lengths of caudal-fin rays to their distal tips. Some specimens with distal tips of caudal-fin rays darkly pigmented.
Blind side uniformly yellowish-white; some specimens with a few, small, irregularly-scattered, reddish-black, pepperdot-size melanophores on some scales (best observed under high magnification); when present, pepperdots primarily on scales overlying pterygiophore regions of dorsal and anal fins. Some specimens also with dark patches of melanophores on margins of caudal peduncle. Blind sides of dorsal and anal fins without pigment on anterior fin rays, but with small melanophores on fin rays and membranes in posterior halves of these fins. Blind-side pectoral and pelvic fins off-white. Blind side of caudal fin with similar pigment to that on ocular side.
Juvenile color patterns ( Figure 6 View FIGURE 6 A–B). Juveniles (to about 60 mm SL) and a few larger specimens sometimes with slightly different color pattern, especially that of ocular side, than those of larger fish. The most conspicuously different pigmentation features of these specimens are three longitudinal series of darker, elliptical to circular blotches. All blotches contain numerous, conspicuous, small melanophores. One series, located medially along lateral line, usually contains 4–6 large (ca. equal to 40% of ED), nearly spherical blotches. A second longitudinal series appearing on the body dorsal to the lateral line and near bases of dorsal- and anal-fin rays consists of 3–5 blotches, while the third longitudinal series (3–4 blotches) lies ventral to the lateral line near bases of the anal-fin rays. Blind sides of juveniles are uniformly whitish, which is similar to that of adult fishes, but lack pepperdot melanophores on their blind-side scales. The peritoneum is dark black and more conspicuous through the juvenile’s thinner abdominal walls. All dorsal- and anal-fin rays are pigmented as those in adults, but differ in their pattern of 2–4 darkly-streaked fin rays alternating with 2–3 more lightly pigmented fin rays. Two black spots feature prominently on caudal fins of juveniles as small as 41 mm SL.
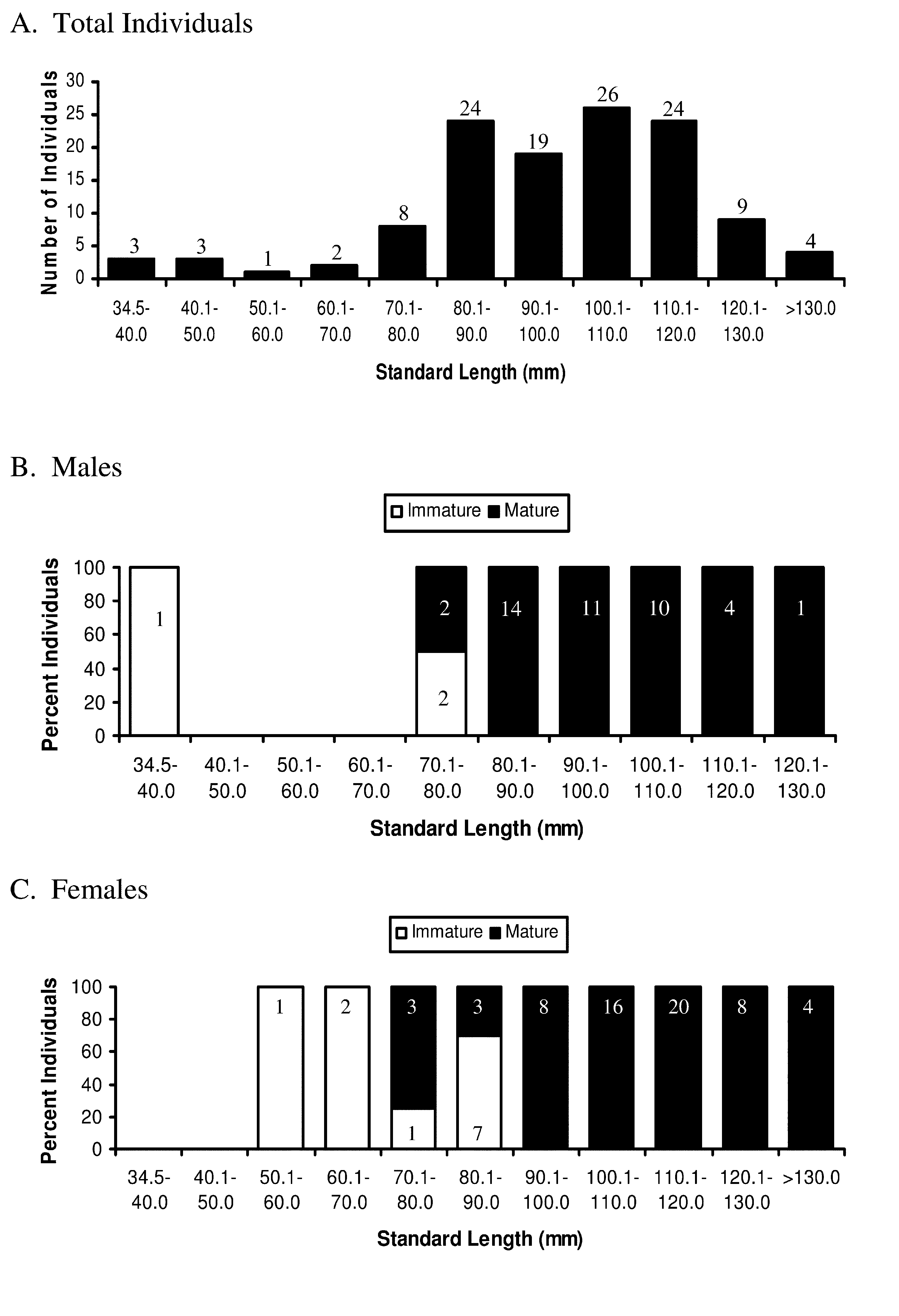
Size and Maturity. To learn about size at maturity, gonads of 124 specimens of A. flemingi , ranging in size from 34.5 to 140.7 mm SL ( Figure 7 View FIGURE 7 A), were examined. Among these fish are 44 males (34.9–128.6 mm SL) and 73 (57.6–140.7 mm SL) females ( Figures 7 View FIGURE 7 B and 7C). Only 5/44 (11.4%) of the males are of sizes greater than 110 mm SL (110.1–128.6 mm SL), while nearly one-half of the females examined (32/73 = 43.5%) are greater than 110.1 mm SL.
Among males, only 3 (34.9–76.9 mm SL) are immature ( Figure 7 View FIGURE 7 B). Two of these immature males are 70–80 mm SL, whereas two others in this size range are mature ( Figure 7 View FIGURE 7 B). Most males (41/44) examined are mature and range in size from 74.8 to 128.6 mm SL. All males> 80 mm SL (N= 39) are mature. Among females ( Figure 7 View FIGURE 7 C), 11/73 (23.3%), ranging in size from 57.6 to 89.9 mm SL, are immature. Among these immature females, three measure <70 mm SL and 7 (of 10 females in this size range) are 70–80 mm SL. Mature females (N= 62) range in size from 78.9 to 140.7 mm SL, with all females (N= 36)> 90 mm SL being mature.
Based on the present study ( Figures 7 View FIGURE 7 B and 7C), females attain slightly larger sizes (maximum ca. 141 mm SL) than do males (maximum ca. 129 mm SL). Males also mature at slightly smaller sizes than those when females become mature. Males begin to reach sexual maturity at sizes of 70–80 mm SL and achieve 100% maturity by 81 mm SL, whereas females begin to reach maturity at slightly larger sizes (ca. 79-90 mm SL) and do not achieve 100% maturity until about 90 mm SL.
Maximum size for A. flemingi examined in the present study is smaller than the maximum size (200 mm; unstated whether SL or TL) reported for this species in literature accounts ( Ayling and Cox 1982; Paul 1986, 2000). No fish of this size were encountered during the present study, where the three largest of 154 A. flemingi measure 140.0– 140.7 mm SL (168.9–174.2 mm TL). Nielsen (1961) and Amaoka (1990) also reported much smaller sizes (86–101 and 83–106 mm SL, respectively) for three and four specimens they measured. Although not stated, the 200-mm maximum size reported in earlier literature accounts for A. flemingi most likely reflects a total length measurement. No specimens collected in New Zealand waters are cited to vouch for this size record, and it is unclear if a specimen of Azygopus of this size was actually measured, or if the 200-mm size so frequently reported is based on an estimated size that has been repeated throughout those references. Possibly, this reported length derives from literature ( Norman 1926; Norman 1934; Gomon 2008) recording a 200-mm maximum total length based on specimens of A. pinnifasciatus from Australian waters.
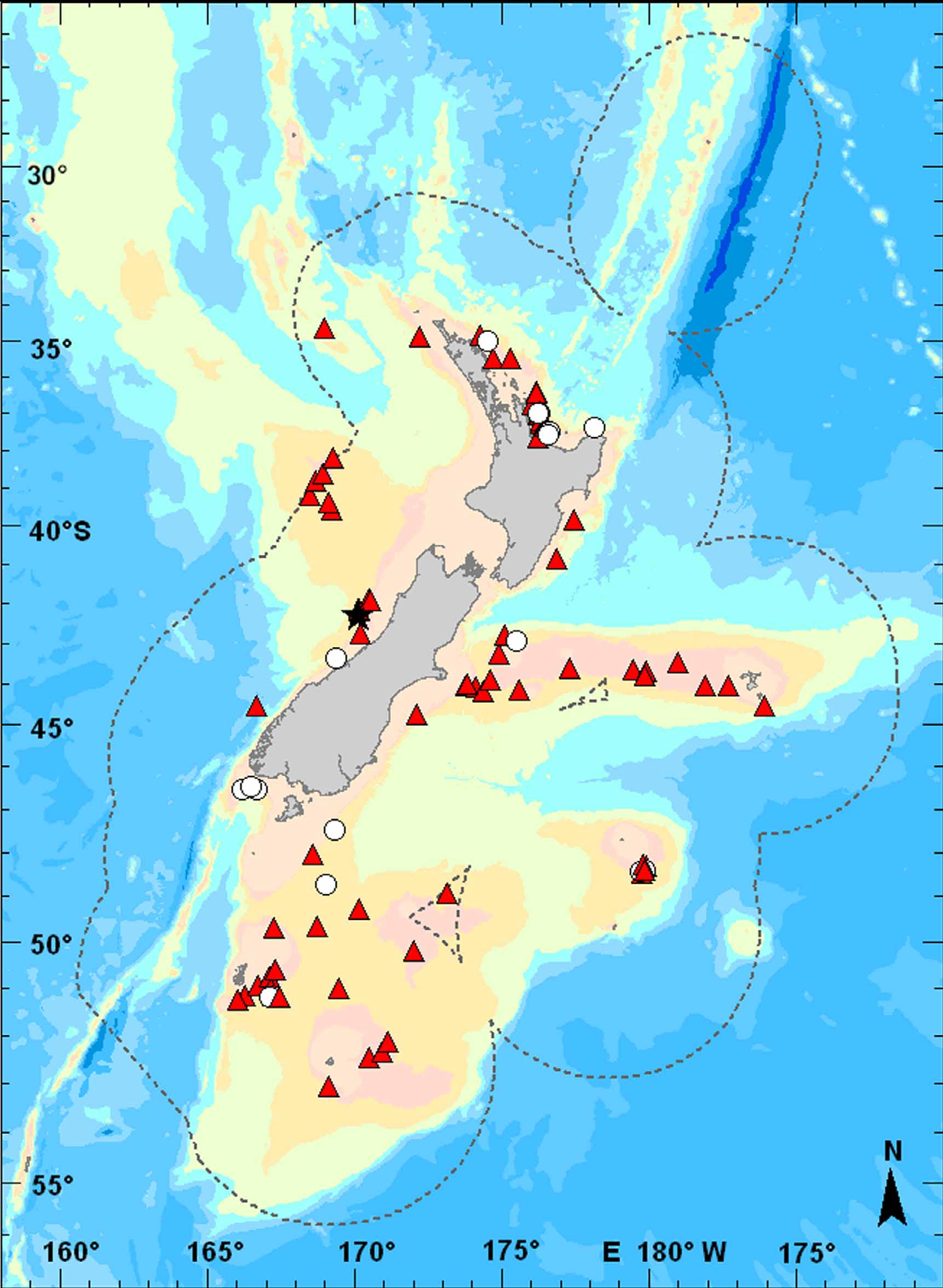
Geographic distribution. Azygopus flemingi is endemic to, and widespread in, deep waters (200–940 m, see below) around New Zealand ( Figure 8 View FIGURE 8 ). Based on museum specimens examined, this species has been captured over nearly 25º of latitude from as far north as the southern West Norfolk Ridge (34º37.44'S, 168º57.57'E) to as far south as the Campbell Island Rise, south of Campbell Island (53º4.00'S, 169º8.00'E). In northern parts of its geographic range, captures include sites off the Poor Knights islands, on the southern Coleville Ridge, as well as several locations off the coasts of Northland and Coromandel. Azygopus flemingi is also known from many locations throughout the southern part of its geographic range including the Hokutika Canyons in the Tasman Bight, off the Auckland Islands, on the Challenger Plateau, the Pukaki Rise and the Campbell Plateau, off Stewart Island, and in the head of the Northern Solander Trough (see also Anderson et al. 1998). Other locations where this species has been taken include the Challenger Plateau, and several museum collections (this study) and other reports ( Moreland 1957; Anderson et al. 1998) also record A. flemingi from deepwater sites off the Chatham Islands and along the Chatham Rise.
Relative Abundance. Studies reporting on fishes taken in trawl catches during expeditions conducted in New Zealand waters ( Nielsen 1961; Amaoka 1990), as well as, ecological works summarizing fish communities around New Zealand ( Shuntov 1979; McClatchie et al. 1997; Anderson et al. 1998; Jacob et al. 1998) record relatively few catches and general low relative abundance of A. flemingi (as A. pinnifasciatus ) in their studies. Anderson et al. (1998), for example, in their comprehensive summary of research trawls where demersal fishes were taken around New Zealand list only 20 of 19,232 stations where Azygopus pinnifasciatus (= flemingi ) were captured. Off southern New Zealand, A. flemingi was also listed (as A. pinnifasciatus ) as a minor component of the demersal fish fauna ( Jacob et al. 1998). Here, it occurred in 14% or less of the trawls and constituted only about 0.38% of the total weight of the catches made in that region. Similarly, low numbers per trawl collection are also recorded in the Ministry of Fisheries research trawl database (data extracted by M. Francis, person. comm., 2 May 2011), however, caution is advised when interpreting these results as mesh sizes in some collections were too large to retain small species such as Azygopus (M. Francis, person. comm., 2 May 2011) and their abundance in these catches may not reflect their actual abundance in their habitat.
The number of specimens per collection based on lots examined in the present study (assuming that all specimens captured were retained) also indicates that A. flemingi is not usually taken in great abundance. The majority of collections (61/93 lots = 65.6% of the total lots) contained only single specimens. Another 13 lots contained two specimens each. Only 9 of 91 lots examined contained five or more specimens, with two largest collections containing 13 and 14 specimens, respectively.
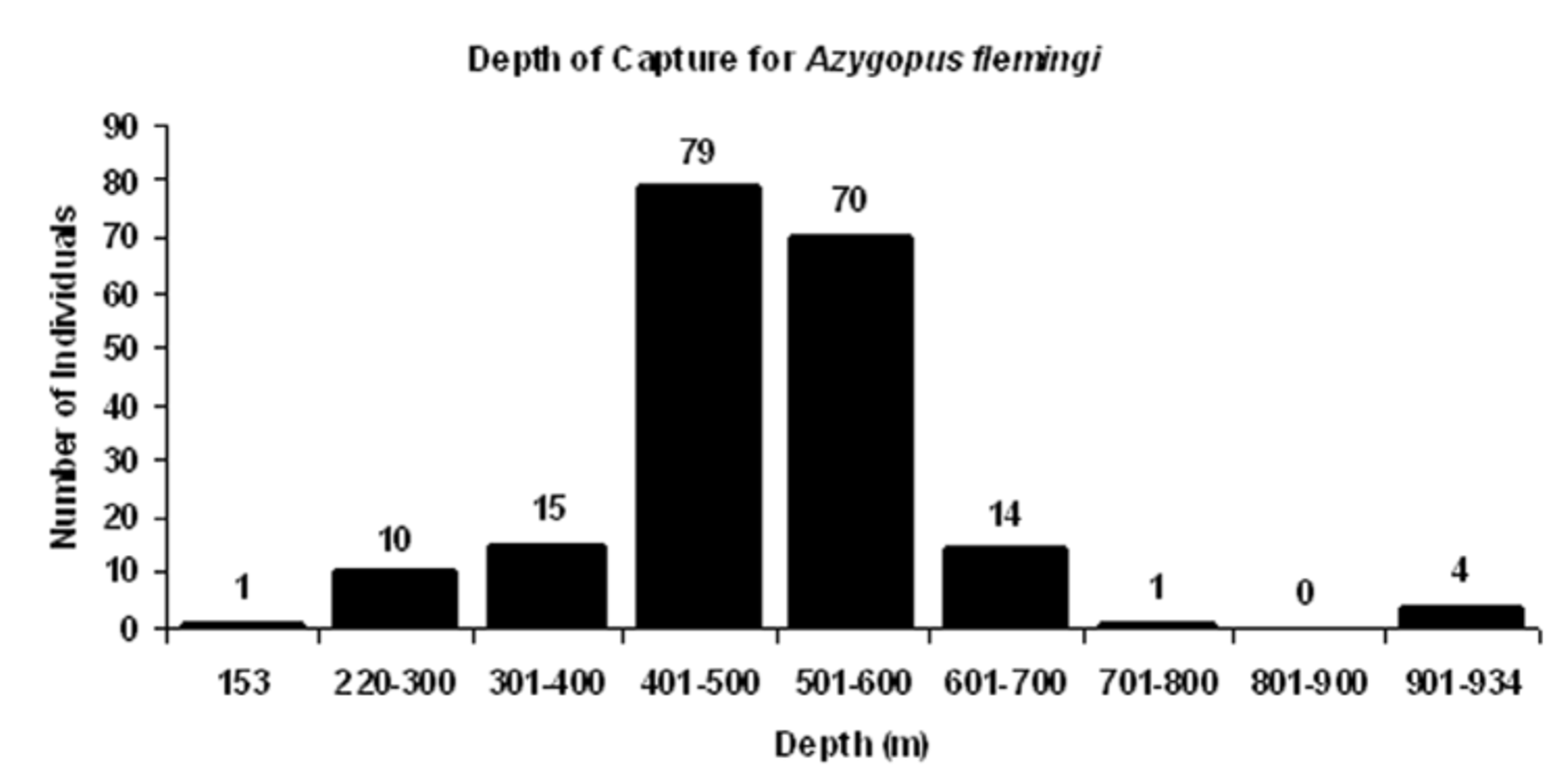
Bathymetric distribution. Specimens of A. flemingi examined in the present study were captured at depths ranging from 153 to 942 m ( Figure 9 View FIGURE 9 ). This depth range is similar to that reported by other authors for New Zealand specimens of Azygopus (listed as A. pinnifasciatus ). Manikiam (1969), for example, reported capture depths of 348–732 m for this species; Shuntov (1979) noted a depth range of 400– 900 m. Ayling and Cox (1982) recorded a depth range of 300–800 m, as did Paul (1986; 2000). Paulin and Stewart (1985) reported a depth range for this species of 200– 600 m. Anderson et al. (1998) recorded a depth range of 200–800 m for Azygopus collected in 20 of 19,232 trawls conducted in New Zealand waters.
The majority of specimens (149/194 = 76.8%) with accompanying depth of capture information were collected between 401–600 m ( Figure 9 View FIGURE 9 ). McClatchie et al. (1997) also recorded a few individuals taken at these depths (550–558 m). Only 11 of 194 specimens in the present study (5.7% of the total) were taken shallower than 300 m; another 15 individuals (7.7%) were taken between 301– 400 m. Relatively few fish (19/194, 9.8% of the total) were caught deeper than 600 m. The deepest captures included 15 fish (7.7%) collected between 601–800 m, and another 4 fish (2.1% of the total) taken in two different collections between 922– 942 m.
Although the smallest fish (34.5 mm SL) was taken at 302 m, eight other small fish (<80 mm SL) examined in the present study were captured at depths ranging from 403 to 556 m, which are depths comparable to those occupied by larger and adult fishes, including some of the largest fishes examined (135–140 mm SL taken at 539– 55 m). Captures of both juveniles and adults at similar depths indicate that these different life history stages occupy similar habitats on the outer continental shelf and upper continental slope.
Remarks. Prior to this study, comparatively little information had been published for A. flemingi or A. pinnifasciatus , especially that dealing with meristic or morphological features. Moreland (1957) reported the first captures of specimens of Azygopus (unknown number) during a survey of fishes from the Chatham Islands area off southeastern New Zealand, but his study did not include any details regarding counts or measurements for these specimens. Some specimens of Azygopus collected during this expedition are curated at NMNZ and were included in the current study. Since Nielsen’s original description of A. flemingi in 1961, only one other study has reported any additional morphological data on this species. Amaoka (1990) identified four specimens of Azygopus taken off New Zealand during cruises of the R/V Shinkai Maru as A. pinnifasciatus . Counts (especially the 47–48 vertebrae; see Table 1), and measurements, as well as color pattern of fish appearing in the photograph accompanying Amaoka’s description are consistent with features of A. flemingi examined in the present study. Several other specimens collected by the R/V Shinkai Maru during these same cruises and now deposited at NMNZ and NSMT are also referred to A. flemingi .
Nielsen (1961), based on three type specimens of A. flemingi from off New Zealand, compared his fish with one paratype and nine other specimens of A. pinnifasciatus taken off Australia. Based on this limited sample size, he noted some general similarities between fish from both locations, but reported several characters that could distinguish specimens from New Zealand waters from those originating in Australian seas. Nielsen reported, for example, that A. flemingi has fewer dorsal-fin rays, fewer ocular-side pelvic-fin rays, more lateral-line scales and more scales above the lateral line, 2–3 fewer gillrakers, a shorter ocular-side maxilla and larger eye diameter, and also noted that his new subspecies differs in the color of its ocular-side dorsal- and anal-fin rays when compared with corresponding features in A. pinnifasciatus . Nielsen also commented that the blind side in both nominal subspecies is uniformly yellowish, but further examination reveals some differences in blind-side coloration between the two species (see below).
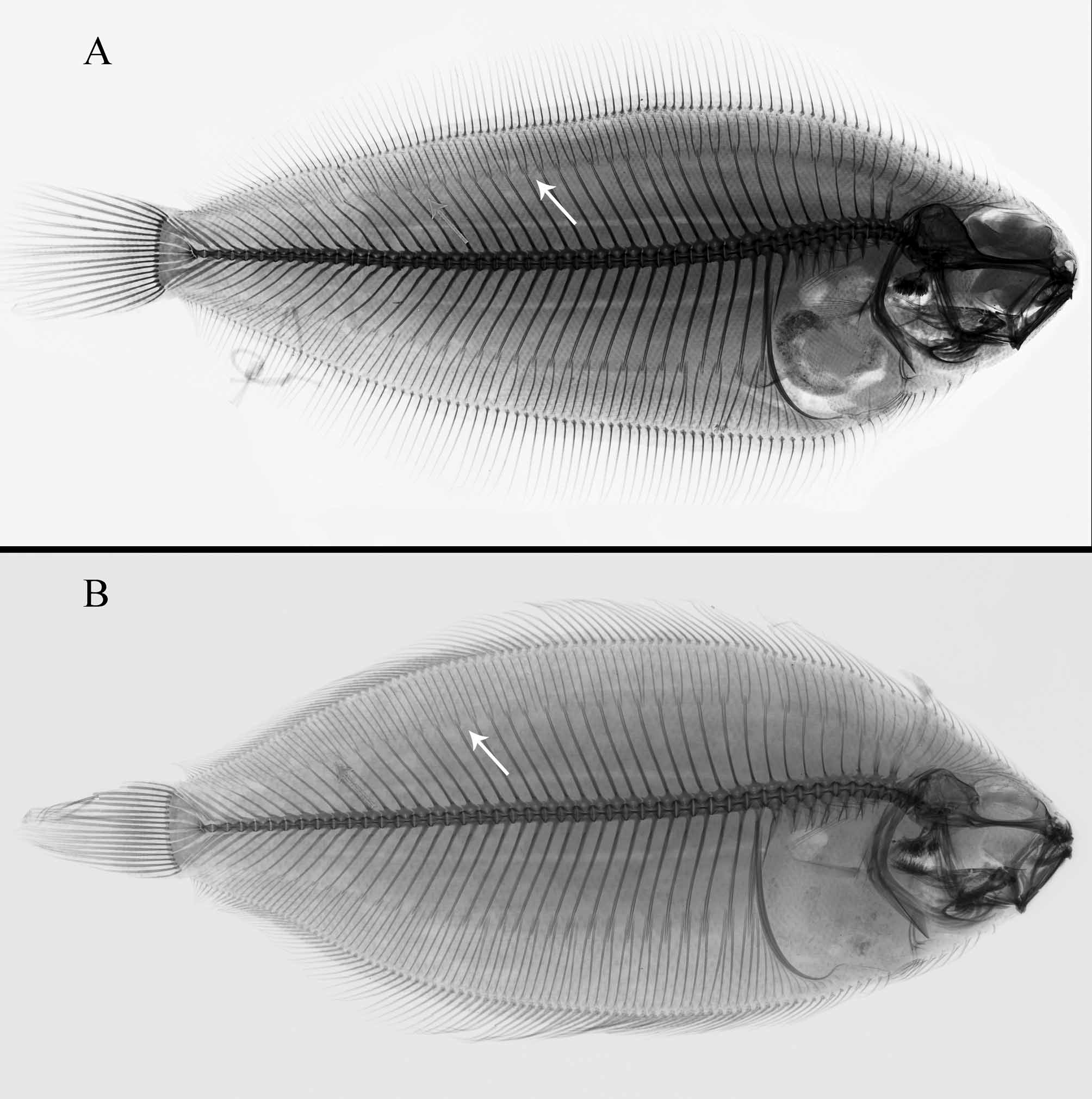
The much larger series of specimens now available of both A. flemingi from off New Zealand and A. pinnifasciatus from Australian seas compared with that accessible to previous researchers provides the opportunity for detailed evaluation of characters historically ( Nielsen 1961) used to distinguish the two taxa, as well as to record new characters found during the present study. Results of this study agree with Nielsen’s (1961) earlier work that A. flemingi is morphologically similar to, but distinct from, A. pinnifasciatus . Considerable overlaps occur in many morphological features examined, several of which are discussed in greater detail below. Among meristic characters, several internal features evident from radiographs also demonstrate close similarity between the species (Table 1). For example, both have similar numbers of supracranial proximal dorsal-fin pterygiophores (6–9 in A. flemingi vs. 7–10 in A. pinnifasciatus ), the same number (1) of proximal dorsal-fin pterygiophores associated with the first neural spine, similar numbers of dorsal-fin pterygiophores inserted into the first and second interneural spaces (2–7, usually 3–5 in the first space in A. flemingi vs. 4–6 in A. pinnifasciatus ) and usually 2–3 pterygiophores inserted into the second interneural space in both species (see Figures 10 View FIGURE 10 A–B). Likewise, the number of anal-fin proximal pterygiophores inserted anterior to the first haemal spine overlap extensively (Table 1) between these two species. Although not diagnostic for distinguishing these species, that both share so many features in common indicates their close relationship.
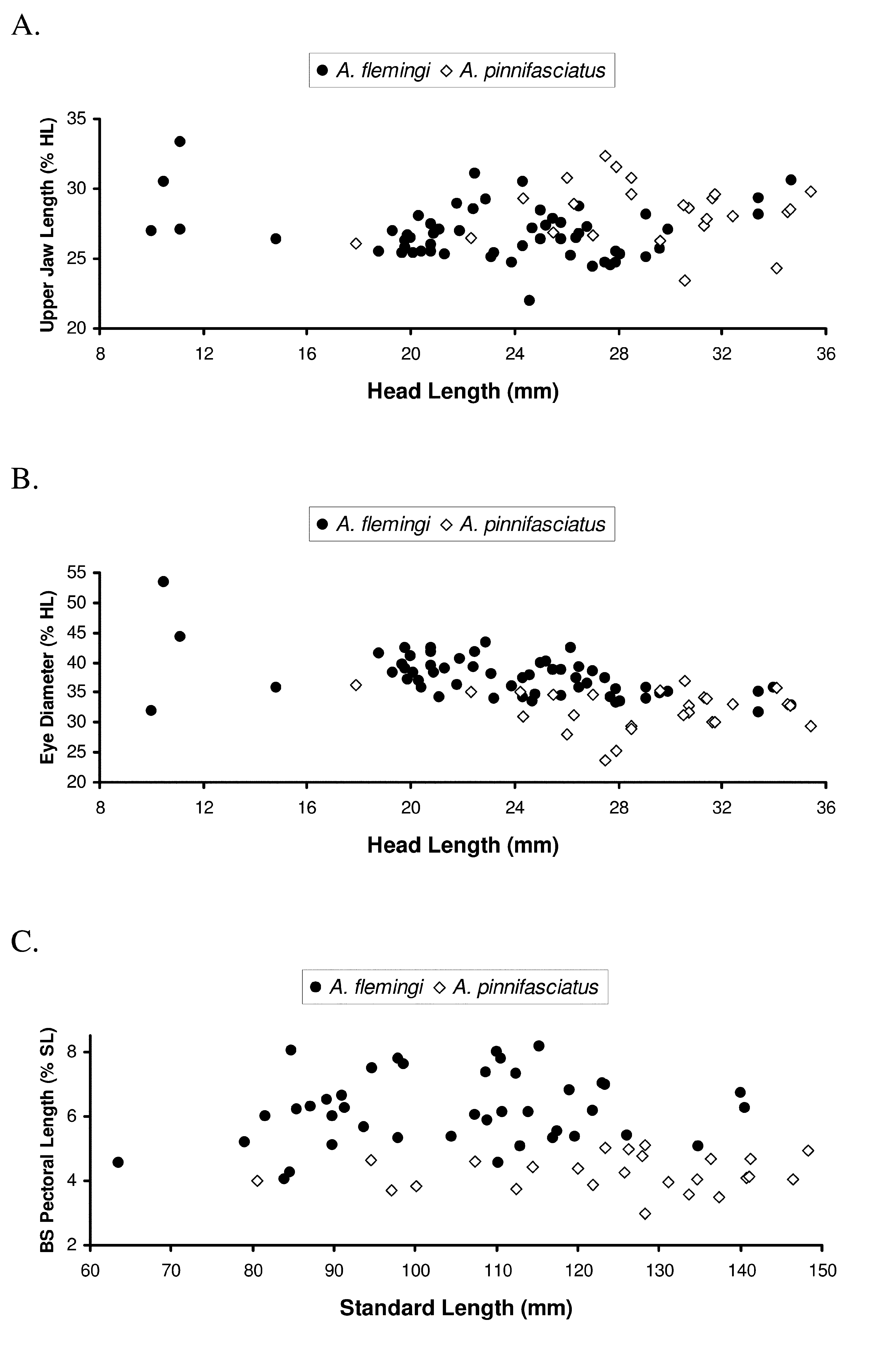
These two species have similar body shapes ( Table 2) as indicated by significant overlaps in several morphometric features including maximum body depth relative to standard length [BD in A. flemingi measures 34.9–48.7% SL ( Nielsen 1961; this study) compared with body depths in A. pinnifasciatus of 38–44% SL (Norman 2006; Gomon 2008) and 36.8–48.0% SL (this study)]. Similarly, the two species have overlapping head lengths [HL 20.8–28.4% SL in A. flemingi ( Nielsen 1961; this study) compared with HL in A. pinnifasciatus equal to 21–24% SL ( Norman 1926; Gomon 2008) and 22.1–26.6% SL (this study)]. Similar levels of overlap between the two species were also found in their postorbital head lengths ( Table 2).
Azygopus flemingi Azygopus pinnifasciatus
All Specimens All Specimens
Holotype N Range Mean SD Holotype N Range Mean SD SIZE (mm SL) 91.0 59 37.2–140.6 99.1 23.29 128.2 24 80.6–148.3 123.0 17.60 BD % SL 38.1 59 34.9–48.7 40.6 2.89 41.2 24 36.8–48.0 42.2 2.36 HL % SL 23.4 59 20.8–28.4 23.9 1.42 23.8 24 21.1–26.6 23.6 1.42 POL % SL 10.9 57 9.6–13.4 11.7 0.85 13.8 24 11.1–14.3 13.0 0.88 BS Pec % SL 6.6 41 4.0–8.2 6.2 1.08 –– 25 3.0–5.1 4.2 0.54 ED % HL 38.9 59 31.7–53.3 38.0 3.73 25.4 24 23.8–37.0 32.1 3.30 UJL % HL 25.3 57 22.0–33.3 26.9 1.96 –– 25 23.5–32.4 28.3 2.11 POL % HL 46.4 57 40.0–58.8 49.2 3.74 57.8 26 47.3–59.9 55.0 2.84 Several characters, originally identified by Nielsen (1961) as useful for distinguishing A. flemingi from A. pinnifasciatus , no longer appear reliable when information from a larger series of specimens is analyzed. For example, Nielsen (1961) reported only slight overlap in numbers of dorsal-fin rays between the two species. Based on data from the present work (Table 1), considerable overlap occurs in this feature and also in numbers of anal-fin rays. Though not completely discrete, counts of dorsal-fin rays for A. flemingi (95–110; Table 1) do range lower than those (102–115) of A. pinnifasciatus ( Norman, 1926; this study). However, overlap occurs throughout most of the mid- to higher end of ranges for dorsal-fin-ray counts of A. flemingi , thus limiting the reliability of this count for identifying only those specimens of A. flemingi and A. pinnifasciatus with the fewest and highest numbers of dorsal-fin rays, respectively.
The length of the ocular-side maxilla was also found not to differ sufficiently enough to be useful for identifying these species ( Figure 11 View FIGURE 11 A). Based on 82 specimens (combined total for both species), this character ranges from 22.0–33.3% HL in A. flemingi , which represent values nearly completely overlapping the entire range of measurements (23.5–32.4.0% HL) for this feature in A. pinnifasciatus ( Norman, 1926; Gomon 2008; this study). Nielsen (1961) reported that A. flemingi has a much larger eye compared with that of A. pinnifasciatus . Larger eye sizes were observed for A. flemingi examined in this study ( Figure 11 View FIGURE 11 B; Table 2), although nearly one-half of the specimens of both species examined (N= 39/83) had overlapping values for this measurement. In A. flemingi , eye diameters range from 31.7 to 53.3% HL (mean = 38.0% HL; Figure 11 View FIGURE 11 B), while those for A. pinnifasciatus range from 23.8 to 37.0% HL (mean = 31.8% HL). Nearly half of the specimens of A. flemingi (N= 28/59) have eye diameters greater than 38% HL, which are larger than any of those observed in the 26 specimens of A. pinnifasciatus measured (largest ED <37.0% HL). Likewise, nearly one-half (11/26=42%) of the A. pinnifasciatus have eye diameters <30% HL, which are smaller than those observed in any of the 59 A. flemingi examined. Although A. flemingi has a larger mean eye size compared with that of A. pinnifasciatus , this character is only reliable for identifying about 52% of the individuals.
Nielsen (1961) indicated that A. flemingi has more lateral-line scales and more scales between the lateral line and highest point of the dorsal profile than are found in A. pinnifasciatus . In the present study (Table 1), A. flemingi was also found to have more lateral-line scales compared with scale counts for A. pinnifasciatus . Again, as for some of the other characters examined, counts of lateral-line scales between these species are not completely different when a larger series of specimens are examined. Nearly one-third (23/76= 30%) of the total number of specimens (both species) have overlapping counts (95–100 scales) for lateral-line scales (Table 1), which restricts the value of this feature in distinguishing the two species. In the few examples where it was possible to make accurate counts, A. flemingi was found to have 3–4 more scales between the lateral line and the highest point on the dorsal profile than was observed in A. pinnifasciatus . However, this count has limited application for identifying specimens because all but a few individuals of both species are missing all or most of their scales, and often these specimens also have many scale pockets torn as well, rendering it difficult to quantify this character accurately. Even when most scale pockets were intact, attempts to estimate scale counts in a subsample of these specimens based only on this count were also unsatisfactory, and consequently it was not possible to clearly distinguish the species.
Other distinguishing characters noted by Nielsen, including numbers of ocular-side pelvic rays and gillraker counts, are more helpful for identifying specimens despite the overlaps present in these features. For example, Nielsen (1961) reported that A. flemingi usually has one less fin ray in its ocular-side pelvic fin. Data from A. flemingi examined in the present study support this conclusion [range in ocular-side pelvic-fin rays = 6–10; most (51/54) with 9 or fewer ocular-side pelvic-fin rays (Table 1) vs. 9–12, usually 10–12 (24/27) ocular-side pelvic-fin rays typically found in A. pinnifasciatus ]. Minimal overlap in this character occurs when specimens (both species) have 10 ocular-side pelvic-fin rays, however, this apparently may happen infrequently as only 3 of 54 A. flemingi examined have 10 ocular-side pelvic-fin rays.
In the original description, Nielsen (1961) also reported that A. flemingi had fewer gillrakers on the lower limb of the anterior gill arch compared with that of A. pinnifasciatus . Results of this study (Table 1) indicate that both numbers of gillrakers on the lower arch and total numbers of gillrakers on the anterior gill arch are more reliable diagnostic characters with little overlap between species. For A. flemingi , counts for gillrakers on the lower limb of the first gill arch are 8–11, with most specimens having fewer than 11 gillrakers, which contrasts to the 10–14 (most with 11 or more) gillrakers on the lower limb of the anterior gill arch found in A. pinnifasciatus . The two species also differ in the total number of gillrakers on the anterior arch, with A. flemingi having fewer (11–16, usually 14 or fewer) compared with that (13–20, most with 15 or more) found in A. pinnifasciatus .
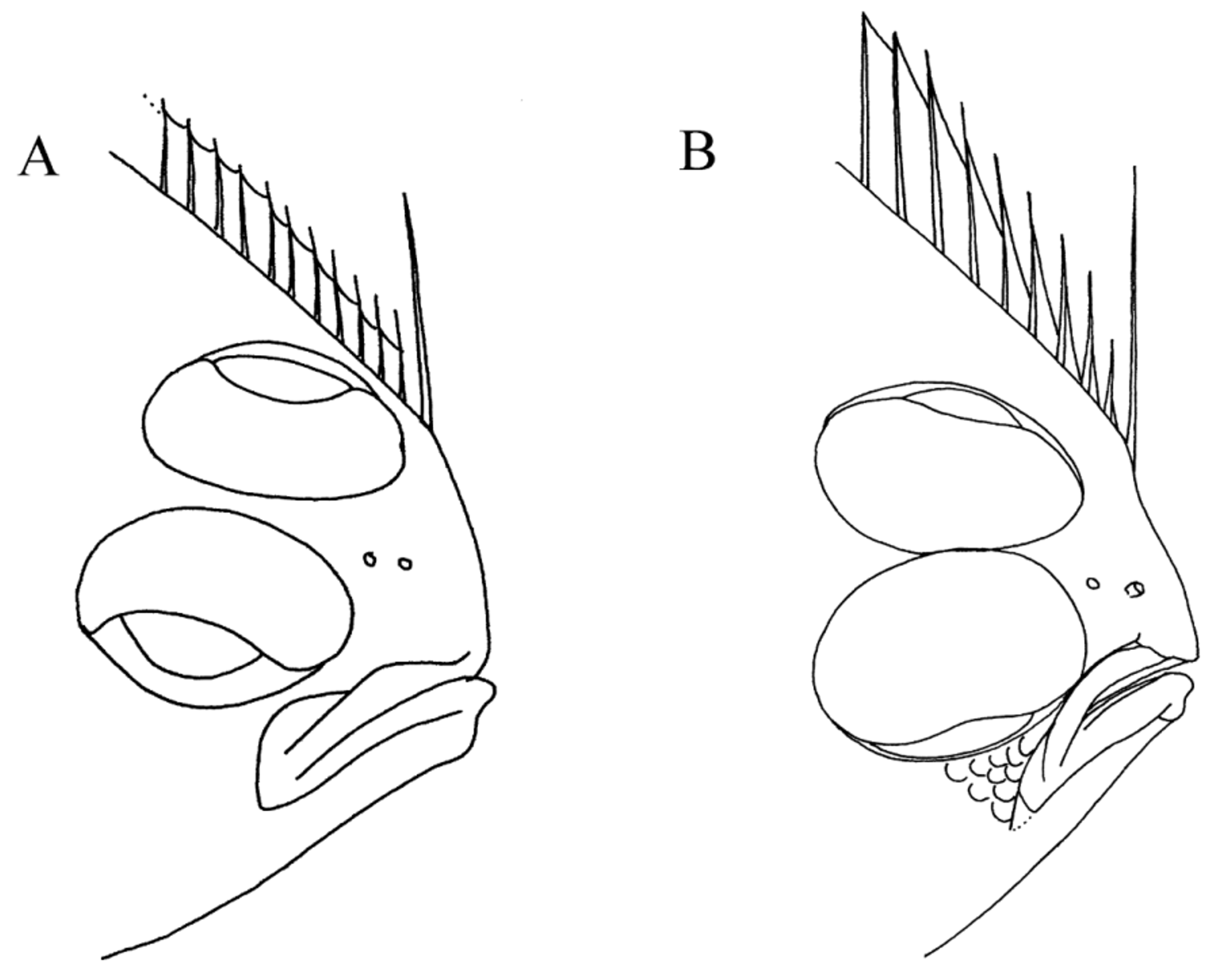
The difference between these species in color pattern of their dorsal- and anal-fin rays, first noted by Nielsen (1961), also appears to be a good character to separate adults of both species, especially when used in combination with other features. Nielsen (1961) noted that A. flemingi has nearly all dorsal- and anal-fin rays pigmented, whereas in A. pinnifasciatus , fin rays in the dorsal and anal fins are distributed as 7–8 groups of 3–5 fin rays streaked with black pigment alternating with groups of ca. 10 unpigmented fin rays (compare Figures 1 and 5 with Figure 12 View FIGURE 12 ). This color distinction is not as obvious in smaller specimens of A. flemingi , which, reminiscent of the color pattern found in adult A. pinnifasciatus , feature small groups of pigmented fin rays alternating with unpigmented (or lightly-pigmented rays).
Several other characters useful for differentiating A. flemingi from A. pinnifasciatus were discovered in the present study. For example, A. flemingi has more vertebrae [46–49; most specimens (46/55) with 47–49 total vertebrae] compared with that found in A. pinnifasciatus , which has 44–46 (usually 45–46; N=26/27) total vertebrae (Table 1). Only seven of 59 A. flemingi had 46 vertebrae. These specimens are discussed in further detail below. Of interest is that in A. flemingi nearly 24% of the specimens have fewer dorsal-fin rays than that found in any of the A. pinnifasciatus examined. Usually in flatfishes (and some other fishes), counts for vertebrae and dorsal-fin rays are positively correlated, so it was somewhat surprising to find that in Azygopus , the species with fewer vertebrae ( A. pinnifasciatus ) typically had higher counts of dorsal- and anal-fin rays. Comparisons of insertion patterns of proximal pterygiophores supporting dorsal- and anal-fin rays (Table 1; see examples illustrated in Figures 10 View FIGURE 10 A–B) reveal how this may have come about. Azygopus pinnifasciatus typically has fewer vertebrae, however, the frequency of interneural and interhaemal spaces where three proximal pterygiophores insert is higher than that found in A. flemingi . Azygopus pinnifasciatus have 8–17 (usually 10 or more; N=18/26) interneural spaces where three pterygiophores insert, while A. flemingi has fewer interneural spaces (3–13, usually 10 or less; N=36/49) where three pterygiophores insert. A similar difference between the two species is also evident in the number of interhaemal spaces with three pterygiophores inserted (7–17, usually 8 or more in A. pinnifasciatus vs. 2–10, usually 8 or fewer spaces in A. flemingi where three pterygiophores insert). Possibly, these data indicate that the number of vertebrae developing in A. pinnifasciatus is reduced from that of the ancestral state, but that fin rays associated with those vertebrae have not been lost (both points evidenced by increased crowding, i.e., greater frequency of three pterygiophores, per interneural and interhaemal space in this species compared with the situation in A. flemingi ).
Other features distinguishing these two species appear in position and angle of the upper jaw relative to the ventral margin of the orbit of the lower eye. In A. flemingi , the upper jaw is angled ventro-posteriorly, with only slight ventral curvature at the distal half of the upper jaw. For most of the anterior two-thirds of its length, the dorsal margin of the upper jaw lies close to, or against, the ventral rim of the lower eye ( Figure 3 View FIGURE 3 A), with little to no space found between these two structures. Since they are so closely juxtaposed, no scales are found in what little space exists between the posterior half of the upper jaw and the ventral rim of the orbit (scales sometimes occur in this space, but only at the very posterior margin of the jaw where a small space occurs between the posterior jaw and ventral orbital rim). In A. pinnifasciatus , the upper jaw is also steeply angled dorso-ventrally, however, in contrast to the situation found in A. flemingi , the distal half of the jaws in A. pinnifasciatus is more strongly curved ventrally ( Figure 3 View FIGURE 3 B). This ventral curvature creates a noticeably wider space between the ventral rim of the lower eye and most of the posterior half of the upper jaw, and in this space there is a small wedge (1–3) of scales.
The two species also differ with respect to aspects of their scale morphology and squamation pattern on the blind side of the body. The most notable difference is evident in the appearance and structure of exposed regions of scales located on the mid-body of their blind sides ( Figures 4 View FIGURE 4 A and 4B). Both species have transforming ctenoid scales ( Roberts, 1993) featuring a posterior region comprising two alternating rows of ctenii marginally, a medial zone formed by 1–2 rows of truncated ctenii, and a basal zone consisting of a calcified plate formed by fused bases of truncated ctenii and subctenii. Exposed portions of blind-side scales in both species feature two distinct regions: (1) the distal region comprising marginal rows of ctenii, a medial zone and calcified plate; and (2) a proximal region lacking ctenii, medial zone, and calcified plate. In A. flemingi , blind-side scales appear to be set closer together because most of the exposed region of these scales (about 70–85%) is covered by the ctenii, medial zone and calcified plate ( Figure 4 View FIGURE 4 A). In contrast, blind-side scales in A. pinnifasciatus appear to be spaced farther apart because distal portions of exposed regions of each scale have a much larger proximal region (i.e., area on scale lacking medial zone and calcified plate) equal to about 60% of the total of the exposed portions on these scales ( Figure 4 View FIGURE 4 B). In A. pinnifasciatus , the distal region of the scale bearing the ctenii, medial zone and calcified plate is only equal to about 40% of the exposed area. Another difference in these species’ scales is that in A. flemingi , no circuli are evident in the very small lateral areas of exposed regions of the scales lacking ctenii, truncated ctenii or the calcified plate. In contrast to this situation in A. flemingi , lateral portions of exposed regions of these scales (areas lacking medial zone or calcified plate) in A. pinnifasciatus are noticeably larger than those of A. flemingi and these areas also feature obvious circuli (see Figure 4 View FIGURE 4 B).
Another difference between these species in their blind-side scales occurs in the extent of the posterior scale margin bearing ctenii. In A. flemingi , about 80–90% of the entire posterior margin of each scale is covered with ctenii. In contrast, in A. pinnifasciatus , ctenii cover only about 50–60% of the central part of the posterior scale margin. These differences in extent of the marginal area bearing ctenii provide a different appearance for the scales in the two species. In A. flemingi , about 80% of the ctenii field of each scale overlaps the ctenii fields on scales located immediately dorsal and ventral to it. In A. pinnifasciatus , a much smaller portion (ca. 30–50%) of the ctenii region of each scale overlaps the ctenii fields of juxtaposed scales located dorsal and ventral to it. The overall visual effect is that, in A. flemingi , ctenii-bearing regions of these blind-side scales are closer set to each other, whereas in A. pinnifasciatus the ctenii-bearing regions of blind-side scales are more widely separated from each other. Azygopus flemingi may also have more ctenii along margins of its blind-side scales compared with those of A. pinnifasciatus , though this needs further verification.
The two species differ also in shape of scale areas containing the medial zone and calcified plate ( Figures 4 View FIGURE 4 A–B). In A. flemingi , the area on the scale where the medial zone and calcified plate are located is crescent-shaped and the posterior margin of the calcified plate has a nearly straight dorsal-ventral profile. In A. pinnifasciatus , in contrast, the scale area with the medial zone and calcified plate is wedge-shaped and the posterior margin of the calcified plate has an obvious anterior extension in its mid-region.
These two species also differ in several aspects of their blind-side pectoral fins. Azygopus flemingi has a longer blind-side pectoral fin (4.0%–8.2% SL; Figure 11 View FIGURE 11 C) compared with that of A. pinnifasciatus (3.0–5.1% SL). Interesting to note is that the holotype of A. pinnifasciatus has only a small remnant of its blind-side pectoral fin. Relative differences in length of the blind-side pectoral fins in these species are also apparent when comparing the posteriormost extent of their longest pectoral-fin rays. In A. flemingi , distal tips of the longest blind-side pectoral rays reach approximately to a vertical through the base of the 10th anal-fin ray, whereas in A. pinnifasciatus , the distal tip of the longest blind-side pectoral-fin ray only reaches to about the vertical through the base of the 5th analfin ray.
Differences also exist between these species in pigmentation on their ocular-side pectoral and caudal fins. In A. flemingi , ocular-side pectoral-fin rays are streaked with darker pigment along their lengths and their distal tips are black, but these fin rays lack banding. In A. pinnifasciatus , the ocular-side pectoral-fin rays also have pigment streaks with darker pigment distally, but in this species the darker pigment on the basal 1/5 of the rays forms an indistinct band, and a second incomplete band is also noticeable near mid-points of these fin rays. The caudal fin of both species features two black spots. In fact, this is the prominent feature of the caudal fins of A. flemingi . Although caudal spots are present in some A. pinnifasciatus , they are often less prominent than those of A. flemingi . Instead, caudal fins of most specimens of A. pinnifasciatus are streaked with dark pigment that obscures any spots which may be present.
Several other characters differ between these species, albeit more subtly, and some of these are more qualitative than quantitative. For example, the symphyseal knob on the lower jaw is more prominent in A. pinnifasciatus than is that of A. flemingi (compare lower jaws of specimens in Figures 5 View FIGURE 5 and 12 View FIGURE 12 ), and A. pinnifasciatus also has a thicker body (not quantified) than that of A. flemingi of comparable size. Although maximum body depth values are similar in both species ( Table 2), the point of greatest depth along the body is different between species (refer to Figures 5 View FIGURE 5 and 12 View FIGURE 12 and Figure 10 View FIGURE 10 ). In A. flemingi , greatest body depth occurs anterior to the body midpoint, usually at about a vertical between anal-fin rays 10–15. In contrast, the point of greatest body depth in A. pinnifasciatus is usually located more posteriorly, at about the body midpoint, or just posterior to that location, but well posterior of a vertical through anal-fin ray 15. Differences between these species are also apparent in their body shapes relative to extent of tapering in body contour posterior to the point of greatest body depth (compare radiographs of specimens of each species, Figure 10 View FIGURE 10 ). In A. pinnifasciatus , posterior tapering occurs gradually over a longer extent of the body compared with the more rapid posterior taper in A. flemingi . Consequently, the posterior body depth is greater in A. pinnifasciatus than it is in A. flemingi . Azygopus flemingi also has a more pointed snout compared with the less pointed, nearly squarish snout of A. pinnifasciatus (see Figures 5 View FIGURE 5 and 12 View FIGURE 12 ).
No previous studies dealing with larger juveniles and adults of A. pinnifasciatus or A. flemingi , collected off Australia and New Zealand, respectively, have commented on the significance of differences in their ocular-side color patterns. In geographically separated populations such as these, it is often difficult to determine whether differences in color patterns are species-specific characteristics, or if they indicate intraspecific variation among populations of the same species. This difficulty is compounded when study subjects are flatfishes and when only small numbers of specimens are available for study. Differences in morphological and meristic characters (discussed above), which are based on a large series of samples, support the hypothesis that A. pinnifasciatus and A. flemingi are distinct species. Likewise, these differences also support the conclusion that ocular- and blind-side color patterns are species-specific diagnostic features for these two members of Azygopus . The ocular-side color pattern of A. flemingi (Figures 1 and 5) typically features numerous, small, irregularly-shaped, dark spots distributed over nearly the entire ocular side. Larger blotches, if present, are hardly visible against the predominant pattern of numerous, smaller, dark spots. Scale pockets of this species are not outlined in dark pigment. In contrast, ocular-side pigmentation of A. pinnifasciatus ( Figure 12 View FIGURE 12 ) lacks the numerous small dark spots found in A. flemingi , instead, this species features fewer, larger, and more diffuse, circular or sometimes ring-shaped blotches, and many scale pockets are distinctly outlined with black pigment. Superficially, both species of Azygopus generally feature uniformly yellowish or whitish blind-side pigmentation (see Figures 1B, 5C and 10C), as had been mentioned by Nielsen (1961). However, when viewed under magnification, pigmentation of blind-side body scales differs between these species. Blind-side scales of A. flemingi , for example, usually lack any small, reddish-brown, pepperdot melanophores. When pepperdots are present in A. flemingi , they occur only sporadically in low densities of one or a few pepperdots per scale ( Figure 4 View FIGURE 4 A), and then only on a small number of scales. In contrast, nearly all blind-side scales of A. pinnifasciatus ( Figure 4 View FIGURE 4 B) are decorated with several to many reddish-brown pepperdot melanophores, and these pepperdots are especially numerous on scales overlying proximal pterygiophores of the dorsal and anal fins. Presence/absence of pepperdots on blind-side scales is a reliable character useful in identifying the two species of Azygopus as these pepperdot melanophores are still evident on blind-side scales of the holotype of A. pinnifasciatus , which was collected nearly 85 years ago.
Comments on specimens with 46 total vertebrae. Nine specimens of A. flemingi , ranging in size from 40.4–116.5 mm SL, have 46 total vertebrae, which is a count more typical of that observed in A. pinnifasciatus . To rule out the possibility that these specimens are A. pinnifasciatus , they were further evaluated. Five of the six larger specimens (84.1–116.5 mm SL) feature ocular- and blind-side pigmentation patterns, caudal-fin coloration and gillraker and lateral-line scale counts consistent with those found in other specimens of A. flemingi . They also lack scales between the lower eye and upper jaw, which is typical of other specimens of A. flemingi . Based on these characters, they were identified with confidence as A. flemingi .
The sixth large specimen ( NMNZ P.044553, 115.9 mm SL, adult female) lacks pigmentation on its ocular side. Its 46 total vertebrae and single scale between ocular-side upper jaw and ventral margin of lower eye are features more similar to those of A. pinnifasciatus . The specimen lacks ocular-side pigment and its blind-side pectoral fin, which hinders identification. However, several characters help to identify this specimen as A. flemingi , including counts of lateral-line scales (113) and gillrakers (9 on lower limb, 13 total), both of which fall within ranges for these features of A. flemingi , as well as its caudal-fin coloration (no streaks), absence of pepperdots on blind-side scales, pointed snout, large eye (35.7% HL), and structure and pattern of blind-side scales, which correspond to features typically found in A. flemingi .
Three smaller specimens (40.4–63.1 mm SL) with 46 total vertebrae have an ocular-side color pattern seen only in juvenile A. flemingi as shown in Figure 6 View FIGURE 6 (no juvenile A. pinnifasciatus examined, so color pattern of this life stage unknown for this species). This consists of irregular to spherical, darker rusty-brown spots covering nearly the entire head and body but also includes a series of larger pigmented blotches. This pattern is reminiscent of the adult color pattern of A. pinnifasciatus . However, none of these three smaller specimens have reddish-brown pepperdots on their blind sides nor do they have dark streaking in their caudal fins, features typically found in A. pinnifasciatus . Two of these have dorsal- (97, 101) and anal-fin ray counts (82, 82) more typical of those found in A. flemingi . Fin-ray counts for the third specimen are inconclusive. Further hindering positive identification of these three specimens is that their scales are either not developed or are missing, preventing use of lateral-line scale counts and blind-side squamation patterns to help identify them. Gillrakers were not fully developed, so accurate counts for this feature were also not available. No scales were observed between the eye and jaw in these specimens, but it is unknown how early in development this character appears in young A. pinnifasciatus . Based on absence of blind-side pepperdots, caudal-in coloration, and fin-ray counts, these specimens are identified as A. flemingi . However, the difficulties identifying these small specimens highlight the fact that more work is needed on juveniles to find features helpful for identifying earlier life-history stages of both species.



| NMNZ |
Museum of New Zealand Te Papa Tongarewa |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Azygopus flemingi Nielsen, 1961
| Munroe, Thomas A. 2012 |
flemingi
| Nielsen 1961 |