
Phacellophora camtschatica
|
publication ID |
https://doi.org/ 10.5281/zenodo.276585 |
|
DOI |
https://doi.org/10.5281/zenodo.6187071 |
|
persistent identifier |
https://treatment.plazi.org/id/039487B4-7B29-FFFB-759D-3231FAFCF800 |
|
treatment provided by |
Plazi |
|
scientific name |
Phacellophora camtschatica |
| status |
|
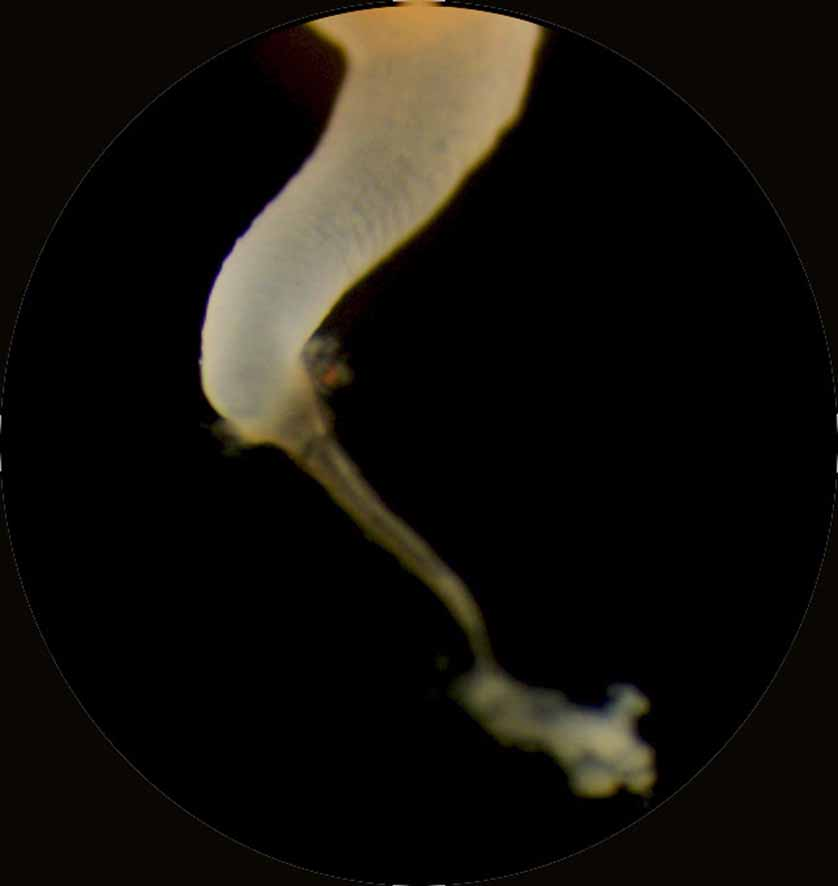
Scyphistoma. Mature scyphistomae (Plate 1, Fig. B) had a mean height of 7.62 mm and a mean oral disc width of 2.45 mm (Table 4c). Tentacles were arranged in a single whorl (ranging from 30–44, mean = 38) around a slightly sunken mouth disc and a cylindrical, four-lipped hypostome. Scyphistomae were yellowishwhite to pale orange in colour. The calyx, perched on a stalk of medium length (44% of TBL), was slender and funnel-shaped. The widest body diameter was the mouth disc (70% of the calyx length). The basal disc and ca. 50% of the stalk were enclosed by a narrow, funnel-shaped periderm tube ( Fig. 4 View FIGURE 4 ) left behind when the scyphistoma changed locations (see below).
Asexual reproduction. This involved typical lateral scyphistoma budding (at calyx and stalk junction, typically one bud per scyphistoma at a time), stolonal budding, and polydisc strobilation (Plate 2, Fig. B; strobilation rates: Table 5).
Two stolonal budding types were observed. In Type 1, thin stolons were produced at the junction of calyx and stalk. One or two new scyphistomae, formed in the middle of the stolon, detached from the stolon after forming a basal disc. In stolonal budding Type 2, a new periderm tube was formed at the tip of the stolon. The old scyphistoma was dragged by the stolon from the old periderm tube, and a tiny tissue residuum remaining in the old tube died after a few days. The stolon was resorbed by the newly located parental scyphistoma.
Ephyra. Newly liberated ephyrae (Plate 3, Fig. B) had a total mean body diameter (TBD), from lappet tip to lappet tip, of 7.77 mm (range: 3.5–10.8 mm, Table 6c) and a mean central disc diameter (CDD) of 5.85 mm, which comprised about 57% of the TBD. Ephyrae had 13–18 marginal arms, flame-shaped and pointed rhopalial lappets, and a single rhopalium per lappet stem, situated between the two rhopalial lappets (Table 6c). Rhopalial lappets measured 53% of the total marginal lappet length (TMLL). The manubrium was of moderate length (≈28% of CDD). There were 1–2 gastric filaments per quadrant in the central stomach. There were 0–2 single tentacle buds above the velar canals. Ephyrae were yellowish in colour with a yellowishorange gastric system; white nematocyst clusters were scattered over the exumbrella.
During subsequent development, velar lappets were not formed but the umbrella rim between the rhopalial lappets extended and grew outwards, forming sinus-like outward curves (pseudo-velar lappets) when the medusa matured.
Development of ephyral gastric system. The gastric system of newly-released ephyrae (Table 3e, Fig. “Stage 1”; Plates 4 and 5, Figs. A) consisted of a central stomach, 13–18 non-forked, arrow-shaped rhopalial canals, and 13–18 very short, non-forked, arrow tip-shaped velar canals. The tips of the rhopalial canals ended at the base of the rhopalium. The tips of the velar canals ended a short distance above the stomach. One or two single tentacle buds were present on the subumbrella above velar canal tips (Plate 4, Figs. B and F).
In the second stage (Table 3e, Fig. “Stage 2”; Plate 4, Figs. D and H) rhopalial canals developed 3–4 pairs of short side branches. The velar canals grew in centrifugal direction. The sides of each velar canal tip developed branches just below its tip (Plate 4, Fig. G; Plate 5, Fig. B) which fused with the first side branches of the rhopalial canals (Plate 4, Fig. G) forming a primary ring canal.
The velar canals and the attached rhopalial side branches grew centrifugally towards the umbrella rim. The primary ring canal morphed into sinus curves (Plate 5, Fig. C). The second pair of rhopalial canal side branches fused with the sinus curves when the velar canal tips reached their level (Table 3e, Fig. “Stage 3”) forming a secondary ring canal.
In the 4th stage (Table 3e, Fig. “Stage 4”; Plate 5, Fig. D–F) the steps of stage 3 were repeated once or twice until a final ring canal was formed below the rhopalia level (Plate 5, Fig. D). The final ring canal produced single tentacles above the velar canal tips (Plate 5, Figs. D and E), and next to the first tentacle buds in a chronological order to form a single row along the ring canal. Additionally, unbranched blind ending canals were produced by the ring canal (Plate 5, Fig. F), and grew in centrifugal direction into the outer region of the marginal lappets. New canal connections formed by "Ausstülpungen" (= diverticula from gastric canals, defined by Stiasny 1921; Uchida 1926), others were disconnected by “Inselbildung” (= island formation, meaning the fusion of the roof and bottom of the gastric cavities, defined by Stiasny 1921; Uchida 1926).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.