
Syngonanthus polyaxis Echtern. & M.T.C.Watan., 2021
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.505.2.5 |
|
DOI |
https://doi.org/10.5281/zenodo.5483852 |
|
persistent identifier |
https://treatment.plazi.org/id/039487CB-AB44-C718-59E9-F9D3FC09FCC8 |
|
treatment provided by |
Marcus |
|
scientific name |
Syngonanthus polyaxis Echtern. & M.T.C.Watan. |
| status |
sp. nov. |
Syngonanthus polyaxis Echtern. & M.T.C.Watan. View in CoL , sp. nov. ( Figs. 1 View FIGURE 1 , 4 View FIGURE 4 , 5 View FIGURE 5 ).
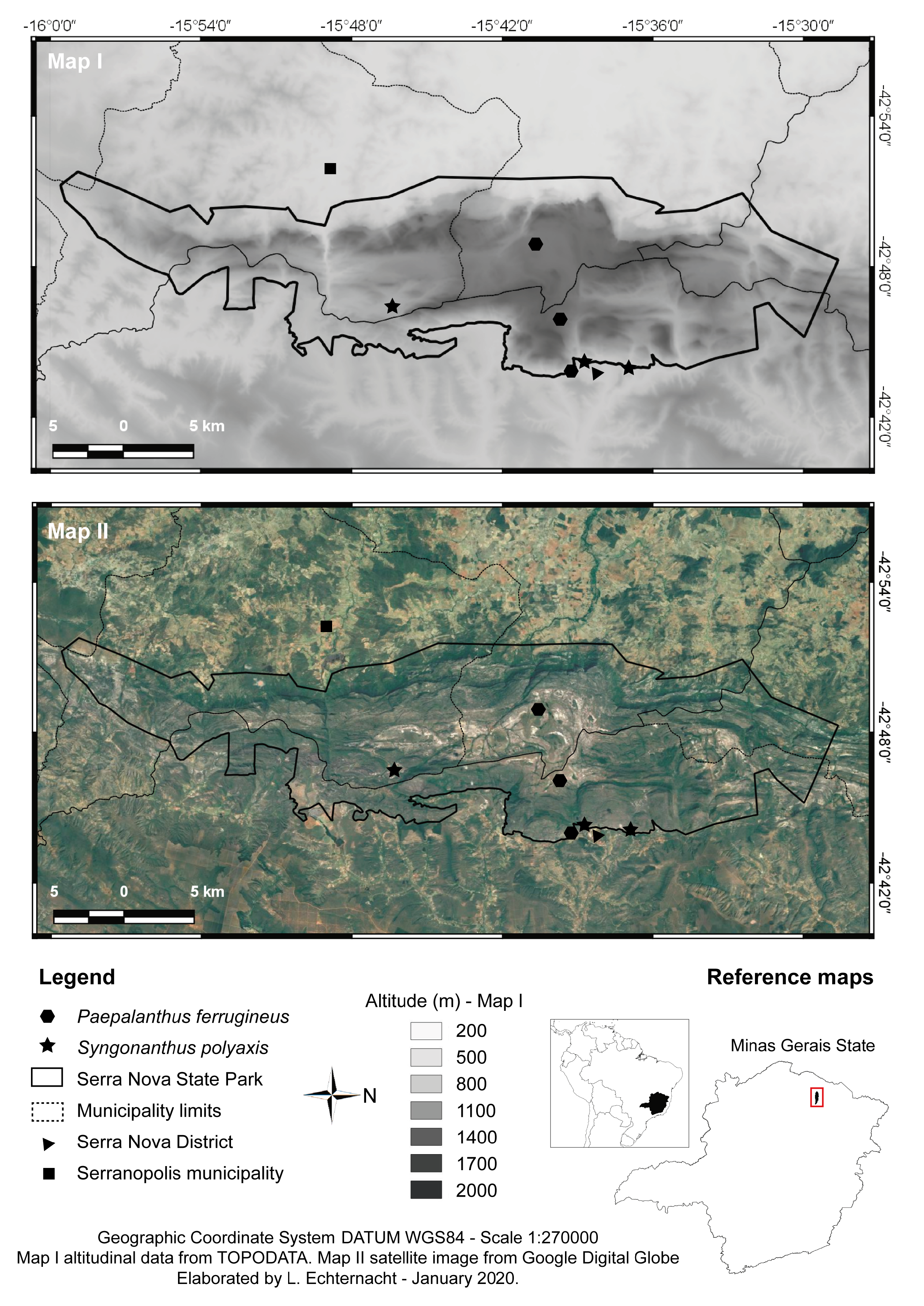
Type:— BRAZIL. Minas Gerais: Rio Pardo de Minas, Parque Estadual da Serra Nova. Alto da serra, passando pela estrada até Gerais Santana , campo rupestre quartzítico, solo arenoso e úmido. 15 o 46’23” S 42 o 46’25” W, 983 m elev., 19 April 2014, L GoogleMaps . Echternacht , T. V . Bastos , M. S . Freitas & A. P. L . Werlang 2405 (holotype OUPR!, isotypes B!, HUFU!, NY!, P!, RB!, SPF!) .
Diagnosis:— Syngonanthus polyaxis is unique within the genus due to its compound inflorescence architecture (synflorescence), with differentiated axes, which a primary axis emerges from the rhizome, bearing deciduous bracts in spiral throughout or rarely naked, with a densely whorl of bracts at apex; followed by one to five thinner secondary axes, naked throughout, with a whorl of less numerous bracts at apex; a tertiary axis is rarely present; the apical part is composed by scapes, bearing capitula, arranged in a terminal umbel-shaped structure. The verticillate bracts approaches S. polyaxis to S. verticillatus Ruhland (1903: 262) and S. graomogolensis Silveira (1928: 342) , however these last two differ from the new species by the undifferentiated synflorescence axis, with more numerous verticilles (3–6 vs 2), and involucral bracts with simple hairs only (vs. simple and capitate).
Perennial herbs, 15–65 cm tall. Roots ca. 1 mm in diameter, cream-colored to orangish, spongy. Rhizome 1–20 × 0.5–1 cm, horizontal to oblique, densely branched, tomentose to villous, with dense, white, curled hairs in young parts, becoming sparsely pilose to glabrescent, with filamentous hairs when older; each branch terminating in a set of overlapping rosettes of leaves, each rosette bearing 1–5 synflorescences. Leaves of the rosette 10–30 × 0.15–0.85 mm, linear to narrowly lanceolate, flat to semi-terete, apex acute to acuminate or acuminate to round, patent to recurved, chartaceous, sometimes striate adaxially with 3 prominent veins; pilose on both surfaces, with simple and capitate hairs, patent, 0.1–0.5 mm long, glabrescent with age; leaf sheath not enlarged. Synflorescence erect, emerging from rhizome, in the leaf axils, with polymorphic axes: a primary axis bearing 1–5 secondary axes, these are sustaining an umbel-shaped structure of 5–30 scapes. Primary axis 4–20 × 0.7–1.8 cm diam, reddish-brown to whitish due to the lanose and deciduous indument, simple filamentous trichomes, up to 1 cm long, glabrescent when older; bracts in spiral along the axis in younger individuals, numerous to few, deciduous, or bracts in spiral absent, 0.3–1 cm long, patent, form and indumentum similar to that of the rosette leaves; numerous bracts densely whorled at the apex, 0.5–2.5 cm long, linear, patent to incurving when young, reflexed with age. Secondary axis thinner than the proximal, 5–20 cm × 0.4–0.9 mm, velutinous and whitish when young, with simple patent hairs, 0.5–1 mm long, soon reddishbrown, pubescent to glabrescent, with simple and capitate hairs; bracts in a whorl restricted to the apex, few, lanceolate to linear, 0.3–1.5 cm long, indumentum like that on the leaves. Rarely a tertiary axis of the synflorescence is present, similar to the secondary axis. Spathes 1.0– 2.8 cm long, chartaceous, cylindrical, lax, obliquely opened, apex acute, erect to recurved, margin entire, pilose abaxially with dense, patent, capitate hairs and less frequently simple hairs, glabrous adaxially. Scapes 4–17 cm long, green to straw-colored with age, densely pilose with appressed, creamcolored, simple hairs and patent, yellowish, capitate hairs. Capitula 3–10 mm diam, obconic. Involucral bracts in 3–5 series, pilose abaxially with capitate hairs more frequent on the external series, filamentous hairs more frequent on the internal series, ciliate, glabrous adaxially, apex acute; external series 1–3 × 0.5 mm, narrowly triangular to lanceolate, stramineous throughout with longitudinal middle band, margins cream-colored to hyaline; internal series 2.5–3.5 × 0.7 mm, slightly oblanceolate, cream-colored and hyaline with a stramineous longitudinal middle band, equaling the flower disc height. Floral bracts absent. Flowers 3-merous, 20–50 per capitulum, the staminate and pistillate flowers generally equal in number, mixed, cream-colored, hyaline when wet, membranaceous. Staminate flowers slightly zygomorphic, 3–3.5 mm long including pedicel of 0.7–1 mm long, pedicel pilose with filamentous hairs; sepals 1.5–2 mm long, shortly fused at base, concave, elliptical to slightly oblanceolate, apex acute, pubescent and ciliate at the upper abaxial surface, tufted, with simple filamentous hairs or glabrescent when older, glabrous adaxially; anthophore ca. 0.5 mm long; petals 2–2.5 mm long, fused, tubular, lobes obtuse, involute after anthesis, glabrous; stamens ca. 3 mm long, epipetalous, anthers dorsifixed, ca. 0.4 mm long, cream-colored to yellowish, introrse, exserted; pistillodes ca. 0.5 mm long, clavate. Pistillate flowers actinomorphic, 2.5–3 mm long including pedicel of 0.2–1 mm in length, pedicel pilose with filamentous hairs; sepals 2–2.3 mm long, free, concave, elliptical, apex obtuse, densely pilose abaxially, ciliate, with simple filamentous hairs, adaxially glabrous; petals 2–2.5 mm long, fused at the upper half, free at base, linear, elliptical, obtrullate or oblong, pilose abaxially with simple hairs; gynoecium 2.5–3 mm long; ovary 0.5–0.7 mm long; style column ca. 0.3 mm long, nectariferous portion 0.5–0.7 mm long, the glandular apex clavate, stigmatic portion simple, 1.5–2 mm long, exserted. Seed 0.5 × 0.2 mm, ellipsoidal, reddish-brown, reticulate, with longitudinal striations more prominent.
Etymology:— The epithet “polyaxis” comes from the Greek, meaning “many axes”, in reference to the different parts composing the synflorescence, a unique characteristic within the genus.
Flowering:— The species was collected in flower and fruit from April to October, during the dry season.
Distribution and habitat:— This species is endemic to the SNSP and occurs in quartzitic Campos Rupestres, growing among sparse, herbaceous to shrubby vegetation. Populations form aggregates and are well distributed within the Park, each composed of tens of individuals, mainly in white-sand soil, which can also appear greyish by the accumulation of organic matter. Plants were observed in depressions in lower plains, where soil becomes soggy during the rainy season.
Conservation status:— Syngonanthus polyaxis is under the same threats as P. ferrugineus , with the aggravation that most of its populations were recorded from the lowlands of the Park, a much more disturbed area, closer to villages, roads and agriculture. In addition, populations are not dense, each being composed of tens of individuals. It is assessed as Critically Endangered following the IUCN (2019) criteria, with an EOO of 3.6 km 2 (B1), an AOO of 3 km 2 (B2), one known location (Ba), and declining quality of habitat (Bb).
Notes and taxonomic affinities:— The species is morphologically similar to Syngonanthus verticillatus and S. grao-mogolensis , both with synflorescence axes bearing whorled bracts throughout. The three species also share abundant capitate hairs on the axis and axial bracts, and similar capitula with lanceolate, stramineous involucral bracts. The axial internodes in S. verticillatus and S. grao-mogolensis species are interpreted as similar parts and not as several, because they look alike, with the same structure, although anatomical comparative data is still lacking. In Syngonanthus polyaxis , the parts differ in width, color, indumentum and bract covering; being the primary axis frequently with densely bracts in spiral, and bearing one to five secondary axes. Additional differences are the number of whorls along the synflorescence (2 [rarely 3] in S. polyaxis , 3–6 in S. verticillatus , and 5–10 in S. grao-mogolensis ) and the presence of capitate hairs in the involucral bracts only in S. polyaxis . The new species also differs from S. grao-mogolensis by densely pilose scapes (vs. glabrous) and petals of pistillate flowers pilose adaxially (vs. glabrous). Comparison among the three species is summarized in Table 2. Syngonanthus verticillatus and S. grao-mogolensis occur in the Espinhaço Range in Minas Gerais as far north as Grão Mogol municipality, around 100 km south of Serra Nova. Sympatric species of Syngonanthus are morphologically distinct and cannot be confounded with the new taxon, although the area being poorly sampled.
Additional specimens examined (paratypes):— BRAZIL. Minas Gerais, Rio Pardo de Minas, Parque Estadual da Serra Nova. Chegada da trilha para o córrego da velha a partir da vila, próximo da Pedreira, 815 m elev., 15 o 36’55” S 42 o 43’59” W, 18 April 2014, L GoogleMaps . Echternacht , T. V . Bastos , M. S . Freitas & A. P. L . Werlang 2403 ( HUFU!, OUPR!); ao lado da sede do IEF, entre vegetação herbácea densa, solo arenoso quartzoso e úmido, 828 m elev., 15 o 38’43” S 42 o 44’12” W, 20 April 2014, L GoogleMaps . Echternacht , T. V . Bastos , M. S . Freitas & A. P. L . Werlang 2426 ( HUFU!, OUPR!); próximo ao córrego da Velha e à Pedreira, no caminho para o Escorregador, 815 m elev., 15 o 36’55” S 42 o 43’59” W, 10 October 2015, L GoogleMaps . Echternacht , M. P . Soares et al. 2640 ( HUFU!, OUPR!)


| L |
Nationaal Herbarium Nederland, Leiden University branch |
| T |
Tavera, Department of Geology and Geophysics |
| V |
Royal British Columbia Museum - Herbarium |
| M |
Botanische Staatssammlung München |
| S |
Department of Botany, Swedish Museum of Natural History |
| A |
Harvard University - Arnold Arboretum |
| P |
Museum National d' Histoire Naturelle, Paris (MNHN) - Vascular Plants |
| OUPR |
Universidade Federal de Ouro Preto, Campus Universitário |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| HUFU |
Universidade Federal de Uberlândia |
| NY |
William and Lynda Steere Herbarium of the New York Botanical Garden |
| RB |
Jardim Botânico do Rio de Janeiro |
| SPF |
Universidade de São Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |