
Hippocampus pontohi, Lourie & Kuiter, 2008
|
publication ID |
https://doi.org/10.11646/zootaxa.1963.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:10371DCA-4076-4F3B-9C43-0E3DBBA6D228 |
|
DOI |
https://doi.org/10.5281/zenodo.5242598 |
|
persistent identifier |
https://treatment.plazi.org/id/853548F1-CEF4-47CD-8A15-7F225B73BCFC |
|
taxon LSID |
lsid:zoobank.org:act:853548F1-CEF4-47CD-8A15-7F225B73BCFC |
|
treatment provided by |
Felipe |
|
scientific name |
Hippocampus pontohi |
| status |
sp. nov. |
Hippocampus pontohi View in CoL sp. nov.
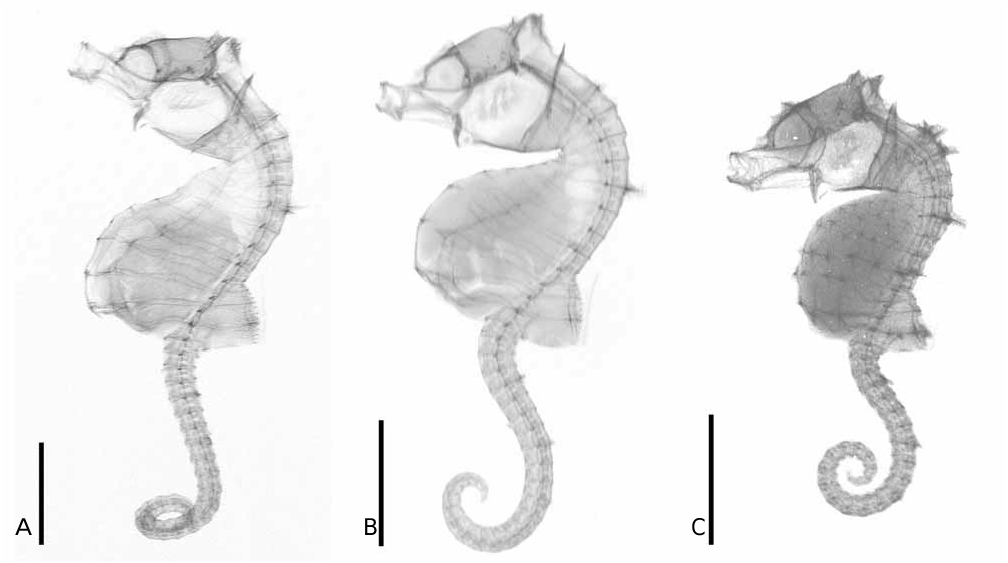
Type material. Holotype: MZB 13593 ( 16.9 mm, male), Lekuan II, Bunaken , North Sulawesi, Indonesia ( 1 o 36'5"N 124 o 45'56"E), 16 m depth, among algae/hydroid crops, 18 July 2003, M. Aw. Figures 2A View FIGURE 2 and 3A View FIGURE 3 . GoogleMaps
Paratypes: MZB 13596 ( 16.8 mm, male), Negeri / Manado Tua, North Sulawesi, Indonesia ( 1 o 37'28"N 124 o 42'54"E), 12 m depth, among byrozoans/hydroids, 22 July 2004 GoogleMaps , R. Lahengko . MZB 13597 ( 16.6 mm, male?), Alung Banua , Bunaken, North Sulawesi, Indonesia ( 1 o 37'7"N 124 o 45'30"E), among byrozoans/ hydroids, 22 July 2004 GoogleMaps , R. Lahengko .
Diagnosis. Extremely small size (height 14 mm, standard length 17 mm); 12 trunk rings; 26–28 tail rings; reduced ossification of inferior and ventral trunk ridges; 14 dorsal fin rays; 9–10 pectoral fin rays; small or absent anal fin; brooding area in males anterior to anus; medium length snout which lacks a bulbous tip; raised, angular coronet; fused (or almost fused in holotype) gill-openings on midline behind coronet supported by raised cleithral girdle; scattered tubercles on trunk and tail; distinctive branching filaments (usually red in life) attached to the enlarged superior trunk ridge spine of the fifth trunk ring; white colour, occasionally with yellow or pink especially along the dorsal surface, with fine red lines on the trunk, and reddish transverse bands across the tail.
Comparison. Hippocampus pontohi is most similar to H. colemani and is primarily separated from the latter species on the basis of tail ring counts ( 26–28 in H. pontohi , versus 28–30 in H. colemani ), raised angular coronet, the following proportions: OD:HL (22.4–26.6% vs 18.6–18.9%), TD9:SL (11.0–15.7% vs 18.9–19.5%) and overall size (SL 16.6–16.9 mm vs 26.5–27.3 mm).
Hippocampus pontohi is much less robust overall than H. bargibanti and lacks the latter’s very large tubercles and bulbous snout tip. It can be further distinguished from H. bargibanti by body colour (white vs purple with red tubercles, or grey with yellow/orange tubercles), fewer tail rings (26–28 vs 31–33), proportionally larger head (HL:SL 20.4–23.9% in H. pontohi vs 14.8–19.5% in H. bargibanti ), less pronounced coronet ( CH:HL 45.1–50.7% vs 46.1–64.1%), narrower snout (SnD:SnL 79.2–87.7% vs 82.2–144.1%), larger orbit (OD:HL 22.4–26.6% vs 14.2–24.0%), smaller post-orbital (PO:HL 49.0–53.5% vs 51.6–63.3%), longer trunk length (TrL:SL 32.4–33.9% vs 25.7–29.5%) and shorter tail length (TaL:SL 43.7–45.8% vs 53.3–57.1%).
It can be distinguished from H. denise by its body colour (white vs orange), deeper head (HD:HL 59.3–61.5% in H. pontohi vs 41.1–55.7% in H. denise ), shorter snout (SnL:HL 22.6–24.4% vs 27.1–38.7%), larger orbit (OD:HL 22.4–26.6% vs 18.8–23.5%), larger post-orbital (PO:HL 49.0–53.5% vs 39.1–45.2%), longer trunk (TrL:SL 32.4–33.9% vs 23.1–31.7%), shorter tail (TaL:SL 43.7–45.8% vs 47.4–57.2%) and deeper body (TD9:SL 11.0–15.7% vs 4.1–15.5%). Both sexes of H. pontohi have rounded trunk profiles in comparison to female H. denise’s narrow trunk. Furthermore, H. pontohi has a more pronounced coronet, fewer tail rings (26–28 vs 27–30) and fewer pectoral fin rays (9–10 vs 10–11).
Hippocampus pontohi can be differentiated from H. minotaur most clearly on the basis of meristic values: TrR 12 vs 8–9, TaR 26–28 vs 41, PF 9–10 vs 11 and DF 14 vs 7–9. It also has a significantly shallower head (HD:HL 59.3–61.5 vs 75.1–80.2%), longer trunk (TrL:SL 32.4–33.9 vs 18.4–24.7%), shorter tail (TaL:SL 43.7–45.8 vs 56.0–66.6%) and longer dorsal fin base (DL:SL 7.3–8.0 vs 1.5–2.4%).
Description. In addition to the characters given in the diagnosis: head length 20.8% (20.3–23.9%) in SL, and head depth 59.3% (60.8%–61.5%) in HL; snout length 24.4% (22.6–22.7%) in HL without bulbous tip, and snout depth 87.7% (79.2–85.5%) in SnL; orbital diameter 22.4% (22.9–26.6%) in HL; post-orbital length 50.9% (49.0–53.5%) in HL; frontal bone strongly raised posteriorly to form a sharply angled coronet ( Figure 2A View FIGURE 2 ); pectoral fin-base raised; pectoral fin rays 10 (9–10).
Trunk rings (TrR) 12, dorsal surface of TrR1 and TrR2 expanded laterally (but without spines); trunk length 33.6% (32.4–33.9%) in SL; trunk depth just anterior to dorsal fin base 15.7% (11.0–13.8%) in SL; dorsal fin base strongly raised and angled with respect to trunk (highest posteriorly); dorsal fin base starting immediately posterior to 9 th trunk ring and ending immediately posterior to 12 th trunk ring (covering 3+0 rings); dorsal fin rays 14; no external pouch visible in males, developing young housed entirely within trunk region anterior to the anus; anal fin not visible in any of the type specimens; first tail ring quadrangular; tail rings 26 (27–28); tail length 45.6% (43.7–45.8%) in SL.
Sexual dimorphism appears to be limited to differences in the genital region: males with vertical pouch slit, females with slightly raised, circular genital opening (see Lourie & Randall, 2003 for diagram).
Body ornamentation: prominent rounded spine above each eye, on midline of snout between eyes, and on either side of head below coronet; shoulder spine at base of pectoral fin; cheek spine; orbital ring with 12 small spines of alternating colours (brown/white); unbranched filament attached to anterior part of coronet in MZB 13596, other specimens without head filaments; greatly enlarged rounded spines on superior ridge of 5 th trunk ring to which distinctive branched red filaments are attached; greatly enlarged rounded spine on superior ridge of 12 th trunk ring and smaller, but still prominent, rounded spines on the lateral ridge of 8 th trunk ring; small, rounded spines on the inferior ridges of 8 th, 9 th, 10 th and 11 th trunk ring (visible as tiny spines in MZB 13596). Enlarged rounded spines on superior ridge of tail align with bands of colour across 5 th, 8 th (or 9 th), 12 th, and 15 th (or 16 th) rings.
Colour in life: white, pinkish or yellowish-white (slightly darker on dorsal surface) with fine red lines tracing the superior trunk ridge and extending ventrally in broad loop around 5 th and 8 th trunk ring; 3–5 red transverse bands across tail that correspond to the position of enlarged superior tail ridge spines; scattered dark markings on head; sporadic dark markings on ventral midline between horizontal trunk ridges; dark patch immediately posterior to anus ( Figure 4A View FIGURE 4 ). Colour in alcohol: pale cream with mid-ventral markings and bands on tail retained but muted. Dorsal and pectoral fins with dark spots in line near dorsal margin ( Figure 3A View FIGURE 3 ).
Discussion. Two other specimens examined: NMV 24979-001 ( 17.3 mm, male; 16.6 mm, female), Milne Bay, Papua New Guinea ( 10 o 20'S 150 o 25'E), 12 m depth, in Halimeda , November 2002, L. Maleta are tentatively identified as H. colemani on the basis of tail ring number, lack of raised coronet, TaL:SL, HD:HL, as well as their geographic locality. The specimens are, however, much smaller than the type specimens of H. colemani and have other proportions that are more similar to H. pontohi sp. nov. than to H. colemani . It is possible that future research will reveal that H. pontohi and H. colemani represent a cline of variation within a single species. In the interests of clarity, and given the present lack of additional data, comparisons in the text descriptions and Tables 1 to 3 are restricted to the type specimens of H. colemani .
Etymology. This species is named in honour of Hence Pontoh, the Indonesian dive guide who first brought these pygmy seahorses to our attention.
Distribution and ecology. Hippocampus pontohi has been observed on the coralline algae Halimeda , as well as on the hydroid Aglaephenia cupressina (Müller and Severns, pers. comm.). Severns noted it particularly in areas where Halimeda is growing on reef walls. It has been recorded at a number of areas in Indonesia (Bunaken, Cape Sri, Sorong, Wakatobi, Lembeh Straits), at depths of between 11–25 m particularly on vertical walls or in rock fissures (Müller, pers. comm.). See figure 5A for map.
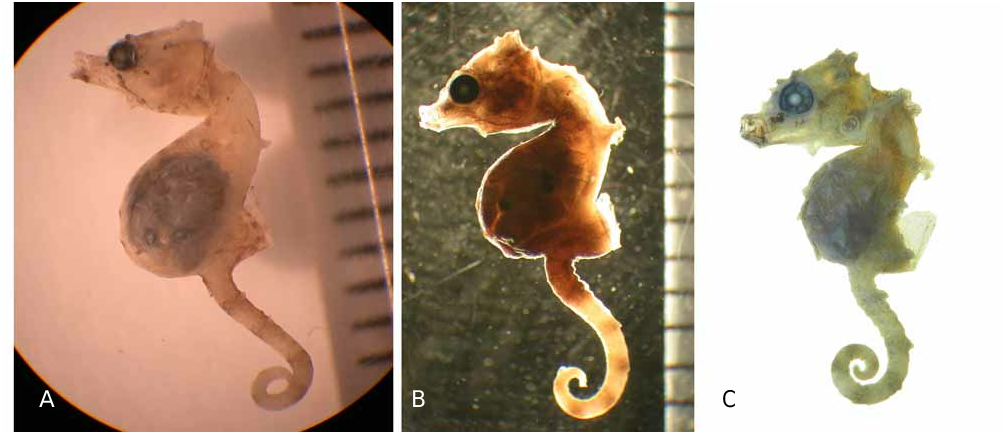
Hippocampus pontohi is commonly found in pairs and, like H. denise , is relatively active (Müller, pers. comm.). Two of the specimens examined were pregnant (MZB 13593 and MZB 13596) and each contained approximately 11 embryos. Both were collected in July.

| MZB |
Museum Zoologicum Bogoriense |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |