
Chilabothrus, 2018
|
publication ID |
https://doi.org/ 10.1093/zoolinnean/zly002 |
|
DOI |
https://doi.org/10.5281/zenodo.5715053 |
|
persistent identifier |
https://treatment.plazi.org/id/0394F107-9437-EB3C-FF74-F91F79D49F11 |
|
treatment provided by |
Carolina |
|
scientific name |
Chilabothrus |
| status |
comb. nov. |
CHILABOTHRUS STANOLSENI View in CoL View at ENA COMB. NOV.
( FIG. 2 View Figure 2 ; TABLE 2 View Table 2 )
Diagnosis: An extinct species of Chilabothrus from the early Miocene of North America, characterized by the following unique vertebral features shared among the referred material: thick zygosphene with a slight prominence on its mid-portion in anterior view; zygosphene roof with a crenate anterior edge, with its median lobe weakly developed and bounded by rounded lateral lobes of the zygosphene articular facets; irregular presence of paracotylar foramina, neural foramina occurring in format of small pits, and variable presence of several foramina distributed in the parazygantral area; neural spine perpendicular to the vertebral centrum, with an elliptical shape in cross-section; articular facet of the zygosphene oval in shape and strongly oriented anterolaterally; vaulted neural arch slightly depressed dorsoventrally in its median portion; deep interzygapophyseal ridges forming a ‘U’ shape; and a shallow posterodorsal notch length that does not exceed more than the half the distance between the pre- and postzygapophyses.
Holotype: MCZ 1977 View Materials partum, an anterior precloacal vertebra.
Referred specimens: Six precloacal vertebrae representing the previous paratypes of MCZ 1977 View Materials (identified by the same catalogue number), comprising five mid-precloacal vertebrae and one anterior precloacal vertebra. MCZ 1978 View Materials , four precloacal vertebrae (same catalogue number), including one mid-precloacal vertebra (previously the holotype of N. barbouri ) and three partial mid-precloacal vertebrae. MCZ 2417 View Materials , three mid-precloacal vertebrae. AMNH FARB 7627 View Materials , two-midprecloacal vertebrae .
Type locality and age: Thomas Farm deposit, Gilchrist County, Florida. Hawthorne Formation. Early Miocene, corresponding to the Early Heminfordian Land Mammal Age (LMA), c. 18.5 Mya. ( Hulbert, 2001; MacFadden, 2001; Steadman, 2008; Beatty, 2010).
Description: All vertebrae of MCZ 1977, MCZ 1978, MCZ 2417 and AMNH FARB 7627 are well preserved and assignable to the same taxon. In anterior view, the zygosphene is thick and wider than the cotyle (zw> ctw). The mid-ventral region of the zygosphene exhibits a convexly projecting border, being more marked in the midprecloacal vertebrae. The prezygapophyses are slightly inclined above the horizontal axis (~10°), showing a higher inclination in the mid-precloacal vertebrae (~13°), whereas they are angled lower in the anterior elements (~10°). A small prezygapophyseal process is observed in some of the vertebrae, extending a little beyond the edge of the articular facet of the prezygapophyses. In the anterior precloacal vertebrae, the dorsoventral height of the neural canal is greater than the transverse width (nch> ncw), whereas the opposite condition is observed in the mid-precloacal elements (ncw> nch). The cotyle is oval in general shape, being slightly flattened dorsoventrally (ctw> cth) in all specimens with the exception of MCZ 1978, in which the cotyle is subrounded (ctw ~ cth). Lateral to the cotyle, there is a paracotylar fossa that in some specimens is associated with paired foramina (i.e. MCZ 1978 and some paratypes of MCZ 1977). The paradiapophyses are strongly oriented lateroventrally with both articular facets clearly differentiated: the diapophyseal articular facet has a strongly projecting convex edge, whereas the parapophyseal articular facet is concave (see the paratypes B, C of MCZ 1977).
Hol. denotes the specimens previously attributed to the holotype material of Neurodromicus ; letters denote an individual vertebra specimen; and dash denotes structure not measured due to the preservation of the specimen. Morphological structures abbreviated in the Anatomical abbreviations’ section.
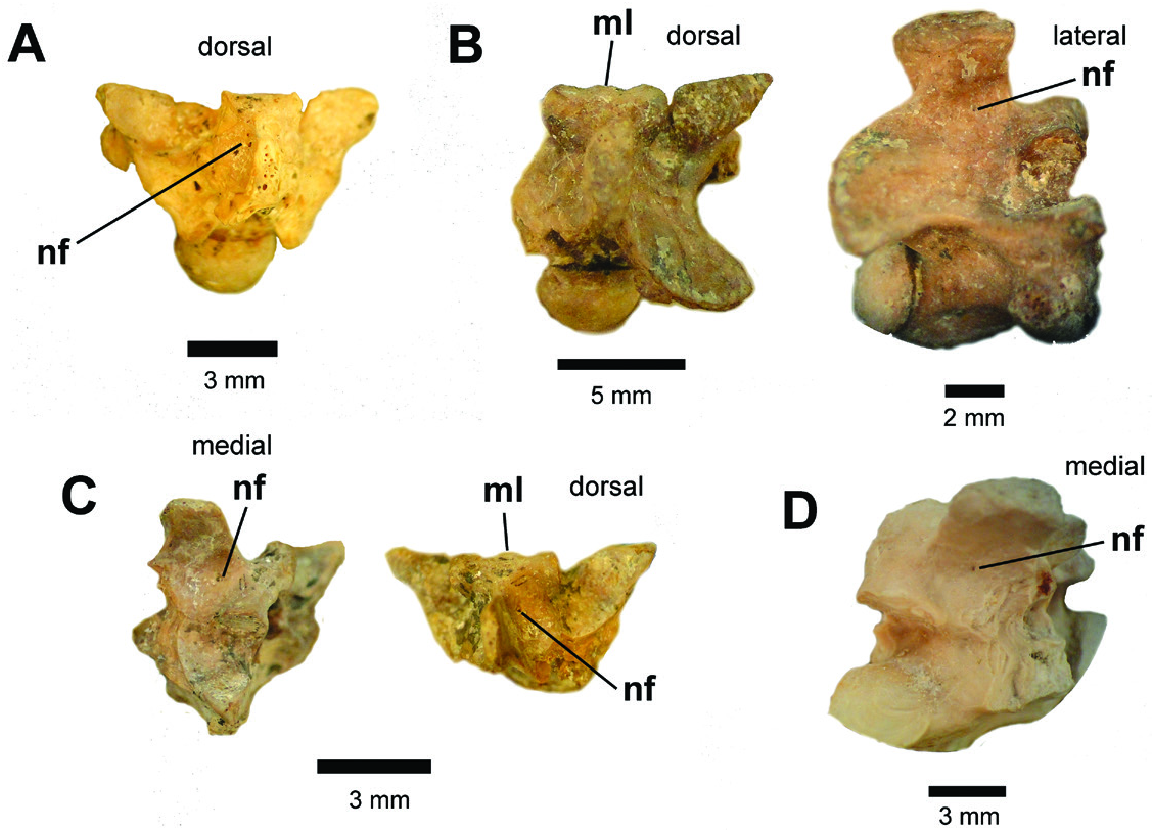
In posterior view, the neural arch is vaulted in all vertebrae. The neural spine of the holotype of MCZ 1977 View Materials is broken at its dorsal tip, whereas a neural spine is not preserved on MCZ 1978 View Materials . In the paratypes C and D of MCZ 1977 View Materials ( Fig. 3 View Figure 3 ) and one specimen of MCZ 2417 View Materials ( Fig. 4F View Figure 4 ), on which the neural spines are entirely preserved, this process is thick and rises from the midportion of the neural arch. The zygantrum is wide, deep and internally bears a set of paired zygantral foramina on each side of the vertebra. External to the zygantrum, several small foramina are present in the form of small pits distributed along the parazygantral area (see holotype of MCZ 1977 View Materials Fig. 2A, B View Figure 2 and paratypes B, D of MCZ 1977 View Materials Fig. 3B, D View Figure 3 ). The postzygapophyses are slightly inclined above the horizontal axis (~10°) in the anterior precloacal vertebrae, whereas they are nearly horizontally oriented in the mid-precloacal specimens. The condyles of all specimens are wider than high (cow> coh), except for one specimen of MCZ 1978 View Materials , which is rounded in shape (cow ~ coh) .
In lateral view, the neural spine is high, anteroposteriorly short and bears a straight dorsal edge. The articular facet of the zygosphene is elliptical in shape and anterolaterally orientated. Lateral foramina are present on the mid-portion of all preserved vertebrae. The posterior region of all centrum is characterized by a marked precondylar constriction. A well-defined haemal keel is present on the ventral surface of the mid-precloacal vertebrae, rising from the ventral margin of the cotyle and extending anteroposteriorly to reach the precondylar constriction, not extending beyond the condyle. The ventral surfaces of the centrum of the precloacal anterior vertebrae do not possess a haemal keel, having instead a hypapophysis. Although broken in the holotype of MCZ 1977 View Materials ( Fig. 2A, B View Figure 2 ) and the paratypes B, D of MCZ 1977 View Materials ( Fig. 3B, D View Figure 3 ), this structure was clearly developed beyond the posterior margin of the condyle .
In dorsal view, the fossils are wider than long (pr–pr> pr–po). The prezygapophyses of the anterior vertebrae are oriented anterolaterally, whereas the mid-precloacal specimens display a nearly transverse orientation. The articular facets of the prezygapophyses are oval-shaped in morphology and longer anteroposteriorly than mediolaterally wide (prl> prw). The anterior edge of the zygosphene displays a crenate morphology, whereby its convex lateral lobes are separated by a smaller median projection (= median lobe sensu Auffenberg, 1963), which does not extend beyond the anterior margin of the laterals lobes. A set of paired parasagittal ridges (sensu Hsiou et al., 2014) extend anteroposteriorly along the roof of the neural arch, beginning on the lateral surfaces of the zygosphene and disappearing just short of the posteriormost edge of the neural arch ( Fig. 2B, D View Figure 2 ). The parasagittal ridges are more pronounced on the mid-precloacal vertebrae than the anterior elements. The neural spine is thick with an ellipsoidal shape in cross-section, although the neural spine of the anterior vertebrae is comparatively thin relative to the mid-precloacal elements. Some specimens possess small neural foramina (= paraespinal foramina sensu Teixeira, 2013) just lateral to the neural spine. These features (similar to the several foramina in the parazygantral area) occur as either small singleton pits or as paired intrusions located on either side of the longitudinal axis of the neural arch, (e.g. the paratype D of MCZ 1977 ( Fig. 3D View Figure 3 ) and paratypes of MCZ 1978 ( Fig. 5 View Figure 5 ). The interzygapophyseal constriction extends from the prezygapophysis toward the postzygapophysis. In the anterior vertebrae, this constriction is relatively anteroposteriorly short, whereas the mid-precloacal vertebrae exhibit a deeper constriction. The posterodorsal notch is relatively shallow in all vertebrae, and does not reach half of the length of the distance between the pre- to postzygapophyses (pnl <50% pr–po).
In ventral view, the length of the vertebral centrum of all specimens is less than the width of the neural arch (cl <naw). In the anterior vertebrae, the hypapophyses rise from the median portion of the centrum, developing anteroposteriorly toward the condyle, extending beyond its ventral rim. In the mid-precloacal vertebrae, the haemal keel originates on the ventral margin of the cotyle, developing longitudinally to reach the prezygapophyseal constriction. The ventral face of the centrum of all specimens has a shallow depression that is more concave in the mid-precloacal vertebrae. The subcentral fossae are delimited laterally by the subcentral margin, being relatively shallow on both the anterior and mid-precloacal vertebrae. The articular facets of the postzygapophyses of the anterior vertebrae are broad and elliptical in outline, whereas they are more subtriangular in shape on the mid-precloacal specimens. In all specimens, the major part of the cotyle is exposed.
Identification and comparisons: All specimens analysed here share with Boinae the following vertebral features: dorsoventrally high, mediolaterally wide and anteroposteriorly short vertebral built; a vaulted neural arch that is wider than the length of the vertebral centrum (naw> cl); well-developed neural spine; thick zygosphene; short prezygapophyseal process; presence of a posterodorsal notch; inclination of the prezygapophyseal articular facets lower than 15°; presence of paracotylar foramina; well-developed precondylar constriction; presence of hypapophyses on precloacal anterior vertebrae; and a haemal keel on mid-precloacal vertebrae ( Rage, 1984, 2001; Lee & Scanlon, 2002; Szyndlar & Rage, 2003; Hsiou & Albino, 2009).
Among the boines, the Thomas Farm fossils specimens strongly differ from the following genera: (a) Eunectes , which presents vertebrae that are markedly larger, wider and more robust; possesses a deeper posterodorsal notch; higher projected parasagittal ridges; and a median tubercle between the neural canal and the zygosphene roof; (b) Corallus , which presents a lower neural spine; higher degree of vaulting of the neural arch; and completely horizontally orientated prezygapophyses; and (c) Boa , which is discussed in greater detail below (see Discussion).
The specimens described herein share with the genus Epicrates the following vertebral features: a thick zygosphene with a protuberance on its mid-portion in anterior view; zygosphene roof with an anterior edge characteristically crenate or entirely straight in dorsal view; irregular presence of paired paracotylar foramina, neural foramina and several foramina occurring in the form of small pits in the parazygantral area; neural spine in elliptical shape in cross-section; articular facet of the zygosphene oval in shape and oriented anterolaterally; vaulted neural arch depressed dorsoventrally in its median portion; interzygapophyseal ridges forming a ‘U’ shape; and shallow posterodorsal notch length that does not exceed more than half of the distance between the pre- and postzygapohyses (pnl <50% pr–po) ( Hsiou & Albino, 2009; Camolez & Zaher, 2010; Hsiou & Albino, 2010; Teixeira, 2013).
Historically, Epicrates was considered a monophyletic genus that shared a sister-group relationship with Eunectes . However, subsequent investigations into the evolutionary relationships of Epicrates have suggested potential paraphyly (based on both molecular and morphological data) with respect to the insular forms from the West Indian island complex ( Epicrates sensu lato) (Sheplan & Shwartz, 1974; Kluge, 1988b, 1989; Burbrink, 2005; Noonan & Chippindale, 2006). Recently, Reynolds et al. (2013) undertook the most comprehensive study of the relationships of Epicrates , proposing the monophyly of the insular West Indian boid forms and formalizing the clade Chilabothrus , which split the insular forms of ‘ Epicrates sensu lato ’ from the mainland ( Epicrates sensu stricto + Eunectes ) and suggested divergent biogegographical and evolutionary histories for these groups.
Despite repeated suggestions of paraphyly, none of the previous works demonstrated explicit morphological differences between the axial skeletons of Epicrates and Chilabothrus . Although at a generic level these genera share a similar combination of vertebral features, there are nonetheless subtle differences that allow for confident referral of the Thomas Farm material to Chilabothrus . These traits can be identified via direct comparison with extant osteological material ( Fig. 6 View Figure 6 ).
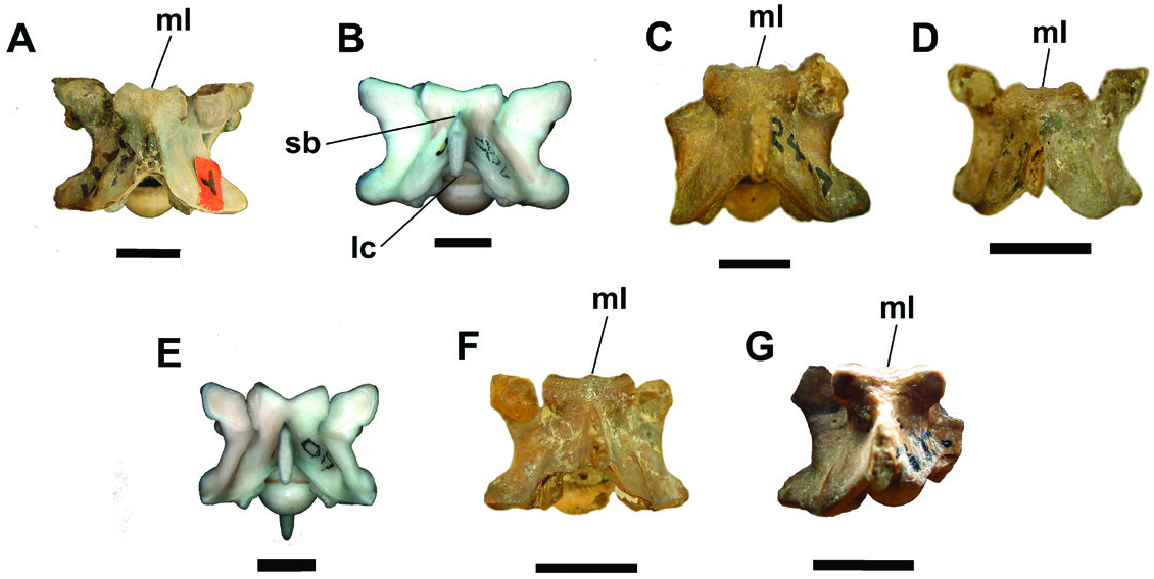
The main differences between these genera can be recognized via the neural spine morphology, the zygosphene and in the consistently greater meanaverage sizes of the vertebral measurements of Chilabothrus ( Table 2 View Table 2 cf. Teixeira, 2013). Epicrates is typified by a high and slender neural spine ( Fig. 4G, H View Figure 4 ), whereas in Chilabothrus the neural spine is low, nearly perpendicular in relation to the vertebral centrum and is more robust ( Fig. 4A–C View Figure 4 ). In dorsal view, both genera display an elliptical-shaped neural spine in cross-section; however, in Chilabothrus the neural spine is proportionally wider than that observed in Epicrates ( Fig. 4I–K View Figure 4 ).
Although the Thomas Farm fossils vary in preservational quality, the completely preserved neural spines of the paratypes of MCZ 1978 ( Fig. 5 View Figure 5 ), MCZ 1977 ( Fig. 3 View Figure 3 ), as well as one of the vertebrae of AMNH FARB 7627, suggests that these fossils share a unique neural spine morphology with extant members of Chilabothrus (compare Fig. 4A–C View Figure 4 with D and F). The neural spines of both groups are nearly perpendicular in relation to the vertebral centrum, shortened anteroposteriorly and appear transversely thickened in dorsal view. These combined neural spine characters differ from the observable morphology of Epicrates ( Fig. 4G, H, K View Figure 4 ).
We also note differences in zygosphene shape variation that are potentially useful for the taxonomic differentiation between Epicrates and Chilabothrus: In dorsal view, the zygosphene of Chilabothrus shows the typical crenate morphology; however, it is worth noting that the median lobe is prominent, broad and characteristically convex ( Fig. 7 View Figure 7 ).
In contrast, the vertebrae of Epicrates show marked variation regarding the development of the median lobe of the zygosphene in dorsal view, ranging from a straight border in which the lobe itself is absent, to the moderately crenate condition and, even, the condition in which the median lobe exceeds the height of the lateral lobes of the articular facets of the zygosphene ( Fig. 7 View Figure 7 ) ( Teixeira, 2013). The morphology of the lateral lobes of the zygosphene articular facets also differs among these two genera, with Epicrates having triangularshaped lateral lobes in dorsal view ( Fig. 7 View Figure 7 ), whereas those of Chilabothrus are rounded ( Fig. 7 View Figure 7 ). There are also some differences in the zygosphene median lobe: in Chilabothrus the morphology of the lobe is characteristically broad and convex, whereas in Epicrates this same process, while also convex, is more ‘triangular’ in shape with a compressed anterior apex (compare the two morphologies in Fig. 7 View Figure 7 ). The Thomas Farm material differs distinctly from these two genera, with the median lobe weakly developed and neither reaching nor exceeding the lateral lobes of the zygosphene articular facets (compare the figures with the extant species: Fig. 6A–C View Figure 6 ; Fig. 4B, C View Figure 4 ; Fig. 7 View Figure 7 ; Fig. 8A, C, D, F, G View Figure 8 ).
In summary, the material from Thomas Farm is assignable to the genus Chilabothrus with respect to the following exclusive combination of vertebral characters: (1) zygosphene with the anterior border crenate in dorsal view with a projection of the median lobe between the rounded lateral lobes of the articular facets of the zygosphene; and (2) neural spine shortened anteroposteriorly, nearly perpendicular in relation to the vertebral centrum, being ellipsoidal shaped and thick. Moreover, here we erect the new combination Chilabothrus stanolseni comb. nov., which represents a hypothetical radiation of extinct species of Chilabothrus into the early Miocene of North America. Additionally, Chilabothrus stanolseni comb. nov. can be identified as a distinct species via a single autapomorphic character: the presence of a weakly developed median lobe, distinct from all conditions observed in the extant genus (see Figs 4 View Figure 4 , 7 View Figure 7 , 8 View Figure 8 ). In this respect, all material previously assigned to the species Pseudoepicrates stanolseni is synonymized with Chilabothrus stanolseni comb. nov.





| AMNH |
American Museum of Natural History |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.