
Malayopotamon weh, Ng, Peter K. L., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3768.5.4 |
|
publication LSID |
lsid:zoobank.org:pub:38F94125-5B01-46B4-BFAA-71A0D9BCFE73 |
|
DOI |
https://doi.org/10.5281/zenodo.6144872 |
|
persistent identifier |
https://treatment.plazi.org/id/039587C0-9E28-F745-FF6D-6560FC381AEE |
|
treatment provided by |
Plazi |
|
scientific name |
Malayopotamon weh |
| status |
sp. nov. |
Malayopotamon weh View in CoL sp. nov.
( Figs. 1–4 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 )
Material examined. Holotype: male (15.5 × 12.1 mm) ( MZB), waterfall, Pulau Weh , 5.82°N 95.28°E, Aceh Province, northern Sumatra, Indonesia, coll. locals, March 2013.
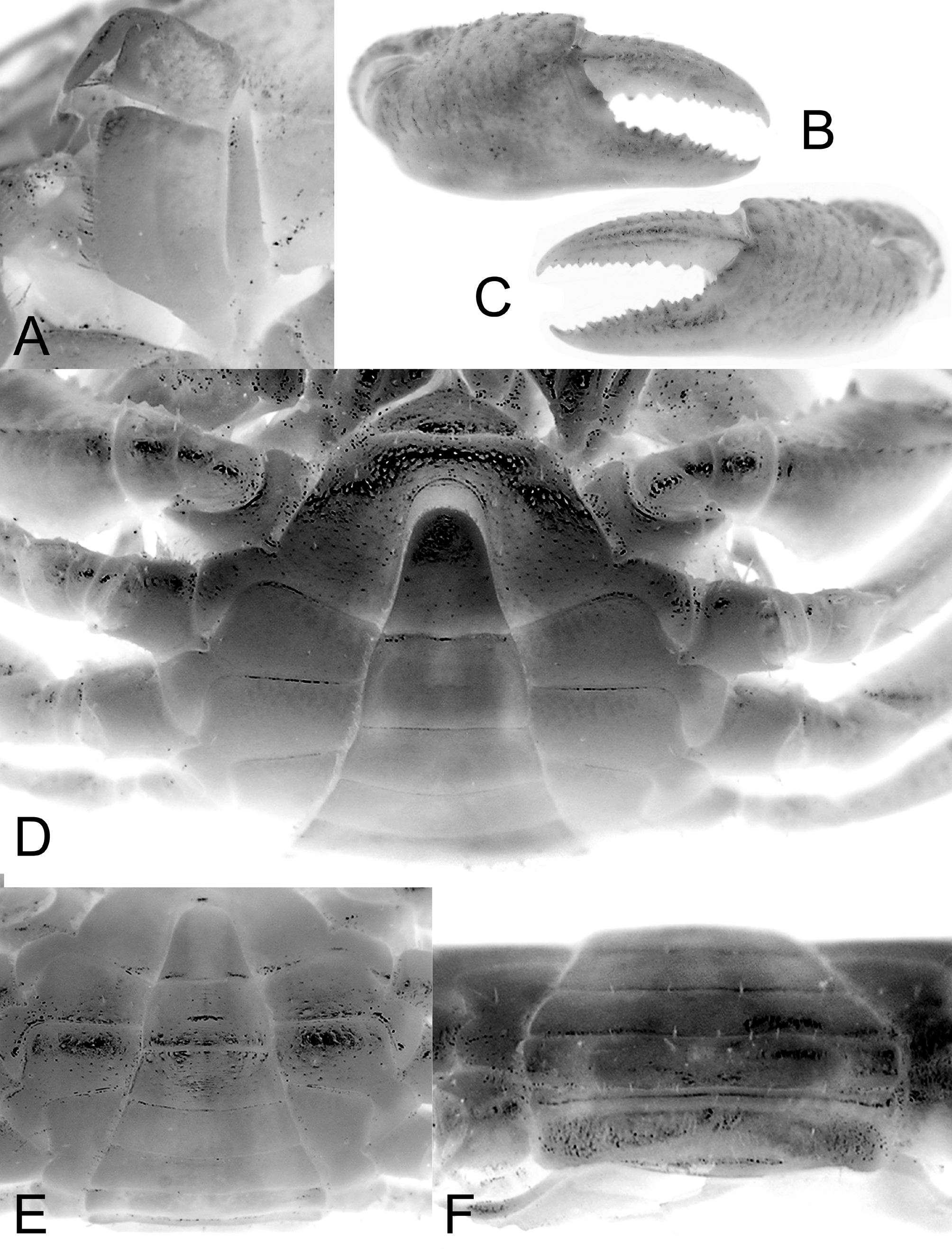
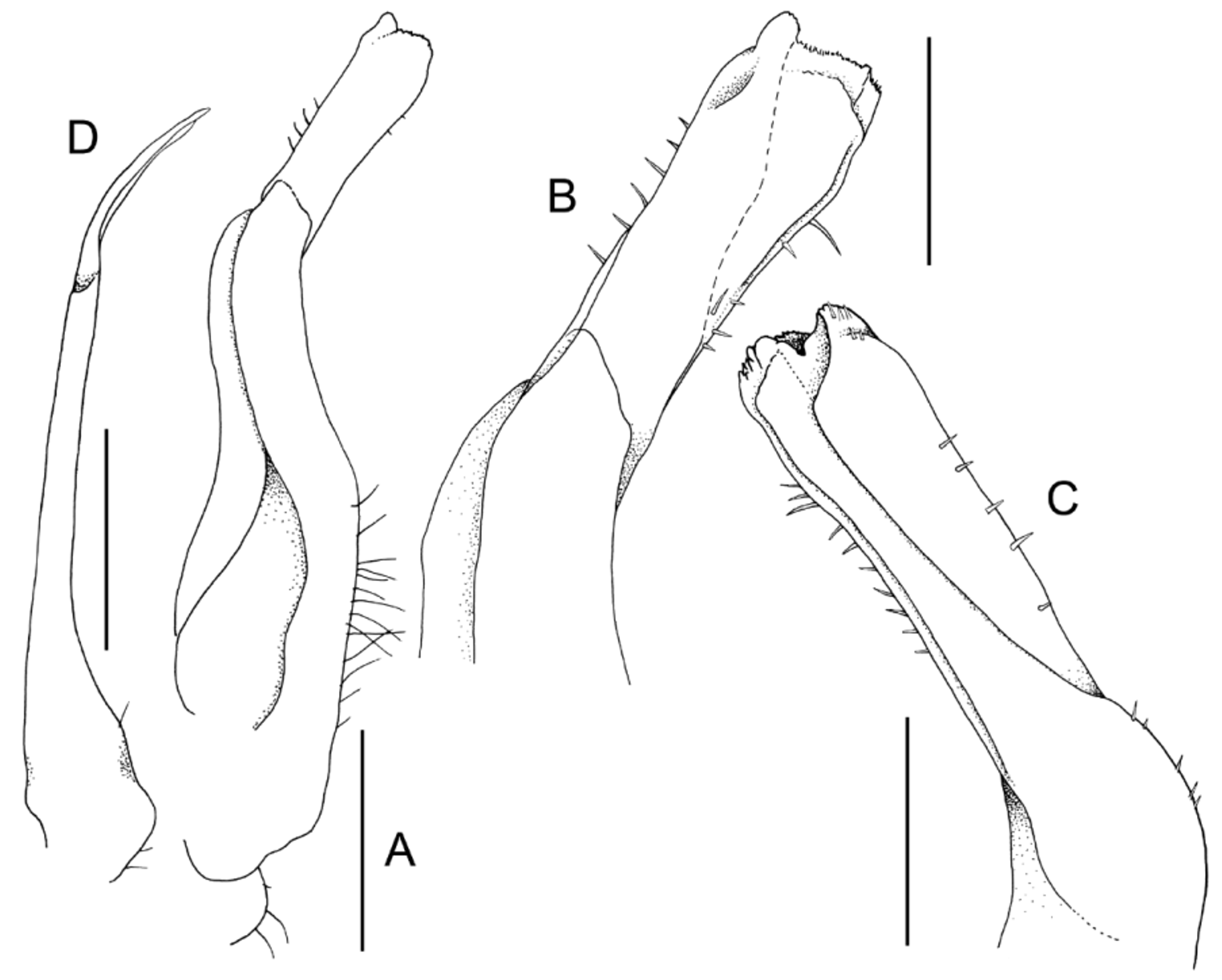
Diagnosis. Carapace with branchial, gastric regions gently convex, not distinctly inflated or swollen ( Fig. 2 View FIGURE 2 A, B); frontal region with low granules; anterolateral regions with distinct low striae, most of branchial regions almost smooth; epibranchial regions with very low striae; mesogastric, urogastric, cardiac, intestinal regions almost smooth, orbital regions almost smooth ( Fig. 2 View FIGURE 2 A); epigastric cristae not clearly demarcated, area marked by rows of transverse striae; postorbital cristae distinct but not sharp crest, demarcated by series of strong, adjacently arranged striae ( Fig. 2 View FIGURE 2 A, B); external orbital tooth broadly triangular, outer margin gently granulated, twice length of inner margin; separated from epibranchial tooth by small notch; confluent with supraorbital margin ( Fig. 2 View FIGURE 2 A, B); epibranchial tooth small, triangular, separated from external orbital tooth by small cleft, not contiguous with postorbital cristae ( Fig. 2 View FIGURE 2 A, B); male thoracic sternum relatively narrow transversely, sternites 1, 2 separated from sternite 3 by deep, sinuous, almost complete suture ( Fig. 3 View FIGURE 3 D); G1 subterminal part gently sinuous with proximal outer margin almost straight; terminal segment about half length of subterminal segment, cylindrical, lateral margins gently diverging to subtruncate tip ( Fig. 4 View FIGURE 4 A–C).
Description of male holotype. Carapace quadrate, smooth, branchial, gastric regions gently convex, not distinctly inflated or swollen ( Fig. 2 View FIGURE 2 A, B). Frontal region with low granules; anterolateral regions with distinct low striae, posterolateral regions with low striae, barely visible; rest of branchial regions almost smooth; epibranchial regions with low striae; mesogastric, urogastric, cardiac, intestinal regions almost smooth, epigastric regions with low transverse striae anteriorly, almost smooth posteriorly; orbital regions almost smooth; suborbital, subhepatic, pterygostomial regions with low granules, striae ( Fig. 2 View FIGURE 2 A). Epigastric cristae not clearly demarcated, area marked by rows of transverse striae, medially separated by distinct Y-shaped furrow, not confluent with postorbital cristae; postorbital cristae distinct but not as distinct sharp crest, discernible by series of strong, adjacently arranged striae ( Fig. 2 View FIGURE 2 A, B). Cervical grooves distinct, relatively broad, joining prominent H-shaped median gastric groove ( Fig. 2 View FIGURE 2 A, B). Frontal margin divided into 2 broad, truncate lobes, separated by shallow concavity; margin of each lobe gently sinuous, separated from supraorbital margin by distinct rounded angle, no lobe discernible ( Fig. 2 View FIGURE 2 A, B). External orbital tooth broadly triangular, outer margin gently granulated, twice length of inner margin; separated from epibranchial tooth by small notch; confluent with supraorbital margin ( Fig. 2 View FIGURE 2 A, B). Epibranchial tooth small, triangular, distinct, separated from external orbital tooth by small cleft, not contiguous with postorbital cristae, transverse striae ( Fig. 2 View FIGURE 2 A, B). Anterolateral margins convex, crested, lined with small sharp granules ( Fig. 2 View FIGURE 2 A, B). Posterolateral margin gently concave, converging towards gently concave posterior carapace margin ( Fig. 2 View FIGURE 2 A, B). Orbits subovate; eye filling up most of orbital space; eye short, peduncle stout; cornea large, pigmented ( Fig. 2 View FIGURE 2 ). Supraorbital margin gently concave, entire, lined with round granules ( Fig. 2 View FIGURE 2 ). Suborbital margin concave, complete, lined with roundgranules ( Fig. 2 View FIGURE 2 C). Posterior margin of epistome gently sinuous; with distinct acutely triangular, narrow median triangle, lateral margins with small submedian fissure ( Fig. 2 View FIGURE 2 C).
Third maxilliped covering most of buccal cavity when closed; ischium subrectangular, with distinct submedian groove which is subparallel to inner margin; merus quadrate, wider than long, surface slightly rugose, anteroexternal angle rounded; exopod relatively slender, reaching to middle of merus, with long flagellum ( Fig. 3 View FIGURE 3 A).
Chelipeds slightly asymmetrical ( Fig. 2 View FIGURE 2 A). Anterior margin of basis-ischium with prominent sharp granules; outer surface of merus covered with distinct transverse striae, inner margins of merus lined sharp granules, upper margin with progressively larger granules with subdistal margin subspiniform; outer surface of carpus distinctly rugose, granular, inner margins distinctly granular, distal angle with short, sharp spine, small basal spiniform tubercle ( Fig. 2 View FIGURE 2 A). Chelae relatively stout; outer surfaces distinctly rugose, granular, especially on upper parts; fingers gently curved, as long as palm; dorsal margin of dactylus gently serrated, outer surface lined with rows of pits; propodal finger with longitudinal groove lined with pits; cutting edges of both fingers with variously sized blade-like teeth, denticles ( Fig. 3 View FIGURE 3 B, C).
Ambulatory legs relatively short, second pair longest, last pair shortest ( Fig. 2 View FIGURE 2 A). Outer surface of merus rugose, dorsal margin uneven, appears gently serrated, subdistal angle distinct but not spiniform; carpus rugose, with dorsal margin cristae, outer surface with distinct submedian crista; outer surface of propodus rugose with low submedian longitudinal ridge, dorsal margin uneven, ventral margin distinctly serrated; dactylus gently curved (almost straight on last ambulatory leg), quadrate in cross section, margins with short, sharp spines, tip corneous ( Fig. 2 View FIGURE 2 A).
Thoracic sternum relatively narrow transversely, surface pitted with lateral parts partially setose, but otherwise smooth, glabrous ( Fig. 3 View FIGURE 3 D). Sternites 1, 2 completely fused to form distinct triangular plate with distinctly convex lateral margins; separated from sternite 3 by deep, sinuous, almost complete suture but lateralmost part distinctly shallow, appears interrupted; sternites 3, 4 completely fused; sternoabdominal cavity reaching to imaginary line connecting anterior edge of coxae of chelipeds ( Fig. 3 View FIGURE 3 D). Male abdominal locking tubercle low, rounded, on median part of sternite 5.
Abdomen triangular, all somites, telson free; somites 3–6 trapezoidal, gradually decreasing in width; somite 6 with length greater than half width, lateral margins gently convex; somites 1, 2 subrectangular, wide, reaching to bases of coxae of fourth ambulatory legs, sternite 8 not visible when abdomen closed ( Fig. 3 View FIGURE 3 D–F).
G1 relatively stout; subterminal part gently sinuous with proximal outer margin almost straight, lined with long setae; terminal segment about half length of subterminal segment, cylindrical, lateral margins gently diverging to subtruncate tip, margin of fold on ventral margin, lateral margins lined with short, simple setae ( Fig. 4 View FIGURE 4 A–C). G2 long, subequal to length of G1; distal segment, about one third length of basal segment ( Fig. 4 View FIGURE 4 D).
Female not known.
Etymology. The species is named after the island where it was collected. The name is used as a noun in apposition.
Colour. The colour in life was orange-brown overall on dorsal surfaces ( Fig. 1 View FIGURE 1 A), with the chelae orange and the third maxillipeds and ventral surfaces pale yellow ( Fig. 1 View FIGURE 1 B).
Remarks. As reviewed by Ng & Tan (1999), species of Malayopotamon can be divided into two morphological groups on the basis of their carapace form (see also Wowor & Tan 2010). Members of one group ( M. batak Ng & Wowor, 1991 , M. granulatum (De Man, 1892) , M. lipkei Wowor & Tan, 2010 ) have the branchial, gastric, orbital and frontal regions strongly rugose and granular, whereas members of the second group ( M. brevimarginatum (De Man, 1892) , M. gestroi (Nobili, 1900) , M. granulosum (Balss, 1937) , M. javanense (Bott, 1968) , M. similis Ng & Tan, 1999 , M. sumatrense (Miers, 1880) , M. tobaense (Bott, 1968) , M. turgeo Ng & Tan, 1999 ) have these carapace regions lightly rugose to almost smooth. The new species belongs to the second group. The G1 structure of M. weh sp. nov. is very distinctive and does not resemble that of any congener, being gently curved outwards, the terminal segment long and cylindrical with the tip subtruncate, and the subterminal segment relatively slender (see discussion in Ng & Tan 1999).
Ng & Tan (1999) discussed the variation in the G1 structure with growth and noted that while it is relatively more slender in smaller males, the general shape and form remains about the same. The degree of bending and/or curvature, and the general structure of the terminal segment has not been observed to vary significantly in species for which there is a good sample size (e.g., M. javanense , M. brevimarginatum , M. granulatum ) (see also Wowor & Tan 2010).
Malayopotamon weh sp. nov. seems to be a relatively small-size species. Although it is only 15 mm in carapace width, a size in which larger species like M. brevimarginatum and M. granulatum are juvenile, the G1 of the holotype of M. weh sp. nov. is relatively well developed, although the reduced setation along the margins suggests it is not fully mature.
Three other species are known from Aceh Province in northern Sumatra, M. batak , M. tobaense and M. turgeo . Malayopotamon weh sp. nov. can immediately be separated from M. batak by its slightly rugose to smooth gastric and cardiac and frontal regions, which in M. batak , is strongly granular and rugose (see Ng & Wowor 1991: pl. 1). The G1 of M. batak has a proportionately shorter and more quadrate terminal segment (Ng & Wowor 1991: fig. 1B–E; Ng & Tan 1999: fig. 2a, b) than in M. weh sp. nov., where the terminal segment is long and cylindrical ( Fig. 4 View FIGURE 4 A–C). Malayopotamon tobaense can be separated from M. weh sp. nov. by the postorbital cristae being confluent with the epibranchial tooth, and the postorbital and epibranchial cristae being more distinct and sharp (see Bott 1970: pl. 58, fig. 89) (postorbital cristae not joining the epibranchial teeth and the postorbital and epigastric cristae not sharp in M. weh n. sp.; Fig. 2 View FIGURE 2 A, B), and the distal part of the subterminal segment of the G1 is distinctly bent with the terminal segment short and quadrate (Ng & Tan 1999: fig. 6) (distal part of the subterminal segment gently curving and the terminal segment elongated and cylindrical in M. weh sp. nov., Fig. 4 View FIGURE 4 A–C). Malayopotamon turgeo is distinctive in having the dorsal surface of the carapace convex, giving it a more inflated physiognomy; and its postorbital cristae is also confluent with and joins the epibranchial tooth (Ng & Tan 1999: fig. 7a) (submarginally interrupted in M. weh sp. nov., Fig. 2 View FIGURE 2 A). The structures of the G1 are also very different. While the terminal segments are both cylindrical, that of M. turgeo is gently curving upwards and the distal part of the subdistal segment is strongly bent (Ng & Tan 1999: fig. 7c, e), whereas the distal part of the subterminal segment is gently curving and the terminal segment evenly cylindrical in M. weh sp. nov. ( Fig. 4 View FIGURE 4 A–C). In addition, while male thoracic sternites 3 and 4 are completely fused in M. weh sp. nov. ( Fig. 3 View FIGURE 3 D), there is a shallow but visible groove present between these sternites in M. batak , M. tobaense and M. turgeo (see Bott 1970: pl. 58 fig. 89; Ng & Wowor 1991: pl. 1; Ng & Tan 1999: fig. 7b).
The small island of Pulau Weh (ca. 156 km 2) is relatively isolated, being about 15 km north of the northernmost tip of Sumatra. Relatively high (about 600 m in altitude), it has several waterfalls and streams. De Man (1895) reported on some intertidal crabs from the island (see also Ng & Liu 1999), but surprisingly, no freshwater decapods have ever been recorded. The present record of a freshwater potamid crab is therefore noteworthy.


| MZB |
Museum Zoologicum Bogoriense |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.