
Gulella deviae, Herbert, 2006
|
publication ID |
https://doi.org/10.1080/00222930600845218 |
|
persistent identifier |
https://treatment.plazi.org/id/039587D1-FF95-FF8A-4781-3908FCA48896 |
|
treatment provided by |
Carolina |
|
scientific name |
Gulella deviae |
| status |
|
Family STREPTAXIDAE Gray, 1860 View in CoL
Gulella deviae sp. n.
( Figures 5–12 View Figures 5–7 View Figures 8–11 View Figure 12 )
Etymology
Named for Dr Devi Stuart-Fox, who has collected much valuable terrestrial molluscan material in South Africa, including specimens of this species.
Diagnosis
Shell very small to minute, cylindrical, smooth and glossy; apertural dentition three-fold: a parietal lamella, a protruding labral tooth, a very weak columella lamella; outer lip strongly concave above protruding labral tooth; whorls roundly rebated below suture; last whorl with an internal tubercle situated ca 0.5 whorl behind aperture; terminal part of body whorl lacking axial ribbing; umbilicus closed.
Description
Shell very small to minute, cylindrical, length 2.5–3.1 mm, width 1.20–1.28 mm; length: width 1.96–2.48 (N 513). Embryonic shell ca 0.78 mm in diameter, comprising just over two whorls, but junction with teleoconch indistinct; smooth and glossy ( Figure 8 View Figures 8–11 ). Teleoconch ( Figures 5–7 View Figures 5–7 ) comprising nearly 5.0 whorls; first whorl convex, but subsequent ones rather more flat-sided, as profile becomes cylindrical; suture narrowly indented, upper portion of abapical whorl roundly rebated (excavated); surface smooth and glossy, sculptured only with weak growth-lines. Peristome interrupted extensively in parietal region where base of preceding whorl is covered by a thin inductural layer; elsewhere margin of peristome slightly thickened and reflected to form a narrowly rounded marginal varix. Parietal lip almost horizontal and aperture thus D-shaped, the flat side situated apically. Apertural dentition three-fold ( Figure 5 View Figures 5–7 ): (1) an obliquely curving parietal lamella which is not fused to the outer lip at its insertion on the base of the penultimate whorl, instead extending well above this, leaving a distinct channel between the parietal lamella and the insertion of outer lip ( Figure 9 View Figures 8–11 ); (2) a strong, ridge-like, mid-labral tooth which projects strongly outward when viewed from the side ( Figures 6 View Figures 5–7 , 9 View Figures 8–11 ); (3) a very low columella lamella, often scarcely evident. In addition to the dentition associated with the aperture, there is a distinct tubercle situated inside the body whorl, just above and to the left of the insertion of the columella lip, ca 0.5 whorl behind aperture ( Figures 7 View Figures 5–7 , 10 View Figures 8–11 ). This is visible externally only in fresh (non-calcined) shells, appearing as an opaque white blotch in the otherwise translucent shell ( Figure 7 View Figures 5–7 ). It develops only at or near maturity, and is not a feature of juvenile shells. Outer lip strongly concave above labral tooth, but lacking an external pit underlying the tooth ( Figure 9 View Figures 8–11 ). Umbilicus completely closed ( Figure 11 View Figures 8–11 ) and with no evidence of crenulation or axial ribbing on base or terminal part of last whorl. Shell translucent, uniformly milky-white when fresh; orange-red dried remains of animal visible internally.
Anatomy
Unknown. Although some of the material discussed was live-collected, it was obtained from dried leaf-litter samples. No specimens preserved in ethanol are available.
Material examined
Type material. Holotype: South Africa, Limpopo, Soutpansberg, Hanglip Forest , 22.99966 u S, 29.88986 u E, afromontane forest, in leaf-litter, leg. J. Swaye, January 2002, length 2.64 mm, width 1.28 mm ( NMSA T2032 About NMSA /W1966) . Paratypes: all NMSA unless otherwise indicated. South Africa: Limpopo: Soutpansberg, Hanglip Forest, 22.99966 u S, 29.88986 u E, afromontane forest, in leaf-litter, leg. J. Swaye, January 2002 (T2033/W2808, two specimens; RMNH 101062 About RMNH , one specimen); Soutpansberg , Hanglip Forest , 23.0 u S, 29.9 u E, afromontane forest, in leaf-litter, leg. A. C. and W. H. van Bruggen, 8 February 1965 (T2112/W3649, three specimens); Soutpansberg , Entabeni , Goedehoop Forest , 23.0833 u S, 30.1167 u E, afromontane forest, in leaf-litter, leg. D. Herbert, 20 November 1997 (T2037/W5661, two specimens); Soutpansberg , Entabeni , Goedehoop Forest , 23.067 u S, 30.121 u E, afromontane forest, in leaf-litter, leg. C. Symes, 30 October 1999 (T 2035/V 7488, two specimens; BMNH 20050567 , one specimen). Mpumalanga : Sabie area , Mount Sheba , 24.9391 u S, 30.7118 u E, afromontane forest, in leaf-litter, leg. A. Moussalli and D. Stuart-Fox, 25 February 2004 (T2034/W2051, one specimen); Lydenberg area , Buffelskloof Forest Reserve , 25.3134 u S, 30.4724 u E, afromontane forest, in leaf-litter, leg. A. Moussalli and D. Stuart-Fox, 17 February 2004 (T2038/W2053, one specimen; NMW.Z.2005.035.00001, one specimen); Waterval Boven area , Tulloch Mohr , Fairy Glen , 25.5596 u S, 30.5833 u E, afromontane forest, in gully with rocky slate substrate, in leaf-litter, leg. A. Moussalli and D. Stuart-Fox, 15 February 2004 (T2039/W2054, three specimens); 10 km south of Malelane , Sherlock Farm , 25 u 35.3509S, 31 u 31.2509E, 780 m a.s.l., indigenous forest, in leaf-litter, leg. J. Horn (L6), 21 November 2004 (T2118/ W3651, one specimen); Nelspruit area , Kaapsehoop, 25.6199 u S, 30.7634 u E, afromontane forest, in leaf-litter, leg. A. Moussalli and D. Stuart-Fox, 12 February 2004 (T2048/ W2111, one specimen) GoogleMaps .
Other material. Barberton Nature Reserve, 25.8594 u S, 31.0366 u E, afromontane forest in gully between pine plantations, in leaf-litter, leg. A. Moussalli and D. Stuart-Fox, 12 February 2004 (NMSA W2115, one juvenile).
Distribution and habitat
Northeastern South Africa, from the Soutpansberg (Limpopo) to the environs of Barberton (Mpumalanga) ( Figure 12 View Figure 12 ); in leaf-litter of afromontane forests along the edge of the northern Drakensberg , at altitudes from 750 to 2000 m .
Remarks
The first specimens of this species collected were provisionally identified as Gulella perspicuaeformis ( Sturany, 1898) , to which it is undoubtedly superficially similar ( Figure 13 View Figures 13, 14 ). That species, however, has a strictly coastal distribution, occurring primarily in dune and coastal lowland forest in southern Mozambique and northern Zululand (Maputaland), at the southern end of the East African coastal plain ( Herbert and Kilburn 2004). It has not been found further than 35 km from the coast or at altitudes greater than 250 m. The collection of specimens, perhaps representing G. perspicuaeformis , in afromontane forests far from the coast and at altitudes in excess of 750 m, was thus unexpected. Subsequent study, however, revealed that this afromontane material in fact differs consistently from G. perspicuaeformis in a number of features and represents a distinct, undescribed species, here described as G. deviae . The most obvious feature distinguishing G. deviae from G. perspicuaeformis is the presence of the internal tubercle in the former. This feature had not initially been observed since the first shells collected were calcined and thus no longer translucent. Only when fresh live-collected material became available did this feature become evident. Additional differences include: (1) the greater degree to which the labral tooth projects beyond the outer lip in G. deviae , such that the portion of this lip apical to the tooth is rendered strongly concave; (2) the consistent presence of axial ribbing, albeit weak, on the terminal part of the last whorl, behind the outer lip in G. perspicuaeformis (absent in G. deviae ) (compare Figures 9 View Figures 8–11 and 14 View Figures 13, 14 ).
In terms of the Gulella subdivisions proposed by Herbert and Kilburn (2004), G. deviae lies with G. perspicuaeformis in Group 2CB (Group 6(ii) of Connolly 1939). Of other species within this group, G. elliptica (Melvill and Ponsonby, 1898) has superficial denticles on the columella lip, G. inhluzaniensis (Burnup, 1914) has subsutural riblets and G. pentheri ( Sturany, 1898) has a more obvious columella lamella and is considerably more elongate. None of these species are known to range north of KwaZulu-Natal. Gulella perspicua (Melvill and Ponsonby, 1893) , with apertural dentition resembling that of G. deviae , occurs like G. deviae in Mpumalanga and Limpopo, and shares with it the unusual, roundly rebated suture, but lacks an internal tubercle. It is also considerably larger (length up to 5 mm), often proportionately broader, sometimes has a basal denticle in the aperture and its outer lip scarcely protrudes in the area of the labral tooth.
The true affinities of G. deviae within the genus Gulella s. l. remain to be established. Although superficially similar to G. perspicuaeformis , the two may not in reality be closely related, having arisen in different faunal assemblages, namely afromontane and tropical/ subtropical coastal belt, respectively. The presence of the internal tubercle inside the last whorl is a feature I have not seen described in other Gulella species, suggesting that G. deviae is rather unusual. Some Gulella species are known to possess internal teeth in juveniles ( Herbert and Kilburn 2004), but these are generally situated close to the growing aperture lip and are laid down and resorbed progressively with growth, and ultimately lost once the adult apertural dentition is formed. The species concerned usually have strong axial ribs and often also a spirally lirate embryonic shell. The internal tubercle found in G. deviae is clearly unrelated.
Conservation
Although described for the first time herein, this is probably a result of the small size of the shell, rather than the narrow range of the species. Indeed its distribution is not particularly restricted, spanning more than 300 km of the northern Drakensberg escarpment. However, it is surprising that the species has not been found, despite relatively good sampling, in the forests of the Wolkberg near Tzaneen, suggesting that it occurs only patchily within its total range. Based on the available evidence G. deviae would probably qualify in the ‘‘vulnerable’’ category, in terms of the IUCN red-listing criteria ( IUCN 2001).
Gulella lindae sp. n.
( Figures 15–23 View Figures 15–18 View Figures 19–22 View Figure 23 )
Etymology
Named for Mrs Linda Davis, collections manager and scientific artist, Department of Mollusca, Natal Museum.
Diagnosis
Shell minute, cylindrical, with short but distinct axial riblets below suture; apertural dentition five-fold: a parietal lamella, two labral teeth, a small basal tooth to left of centre and a strong columella lamella, umbilicus open, its margin crenulate.
Description
Shell minute cylindrical, length 1.77–2.28 mm, width 0.82–0.96 mm, length: width 1.967 – 2.478 (N 550). Embryonic shell domed in specimens from coastal localities ( Figures 15– 17 View Figures 15–18 ), flatter in those from localities further inland ( Figure 18 View Figures 15–18 ); diameter ca 0.58 mm, smooth, comprising approximately two whorls, but its terminal limit frequently not easy to discern ( Figure 21 View Figures 19–22 ). Teleoconch ( Figures 15–18 View Figures 15–18 ) comprising 4.0–4.5 whorls; first whorl strongly convex, subsequent ones less so; as whorls cease to expand outward, shell profile becomes cylindrical; suture shallowly indented; surface mostly smooth and glossy, but with short, distinct subsutural riblets, giving suture a milled appearance. Peristome interrupted extensively in parietal region where base of preceding whorl is covered by a thin inductural layer; elsewhere margin of peristome thickened and reflected to form a relatively strong marginal varix ( Figures 17 View Figures 15–18 , 20 View Figures 19–22 ). Apertural dentition five-fold ( Figure 19 View Figures 19–22 ): (1) a parietal lamella, the outer part of which curves obliquely to merge smoothly with outer lip at its insertion on base of penultimate whorl; (2–3) two mid-labral teeth, the lower one stronger and ridge-like; (4) a small basal tooth to left of centre, peg-like in some samples, obliquely trigonal in others; (5) a columella lamella with a strong tooth (shape somewhat variable), which extends outward for a short distance as a low ridge, but does not reach aperture edge. There may in addition be a more deeply set, low collabral thickening between the lower labral and basal teeth ( Figure 19 View Figures 19–22 ). Labral tooth marked externally by shallow pit behind thickened outer lip; terminal part of last whorl with distinct axial riblets. Umbilicus relatively widely patent ( Figure 20 View Figures 19–22 ), ovoid, with margin crenulated by short axial riblets. Juvenile shells frequently with a small low tubercle inside basal lip, near its junction with columella lip ( Figure 22 View Figures 19–22 ); this laid down and resorbed with growth, and its position relative to aperture edge thus variable; a series of up to three such tubercles may be present at any one time, progressing inward from aperture edge and separated by 0.25–0.5 whorl. Shell almost transparent when fresh; orange-red dried remains of animal visible internally.
Anatomy
Unknown. On account of its minute size, this species has only been obtained from dried leaf-litter samples. Although a number of specimens were evidently alive when collected, none were preserved in ethanol.
Distribution and habitat
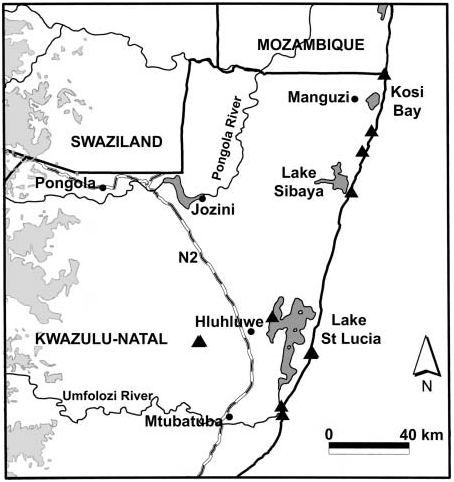
Known only from northeastern KwaZulu-Natal, South Africa ( Figure 23 View Figure 23 ), primarily in the coastal strip from just south of the St Lucia system to Kosi Bay near the border between South Africa and Mozambique, but ranging inland to Hluhluwe-Imfolosi Park in the south; in leaf-litter of coastal and scarp forests, from sea-level to 460 m. Evidently moderately common in dune forests .
Material examined
Type material. Holotype: South Africa , KwaZulu-Natal , Cape Vidal, Bhangazi Hill, 28 u 07.49S, 32 u 33.39E, 60–80 m a.s.l., dune forest, in leaf-litter, leg. Herbert, Seddon and Tattersfield, Site 1, 27 November 1998, length 1.97 mm, width 0.86 mm ( NMSA T 2116/ V 7966) GoogleMaps . Paratypes: all NMSA unless otherwise indicated. South Africa , Kwazulu-Natal : Black Rock , 27 u 07.59S, 32 u 49.59E, coastal forest, in leaf-litter/soil, leg. J. P. Marais, July 2001 (T 2059/V 9295, two specimens); Lake Sibaya area , Lala Nek, 27 u 139S, 32 u 479E, coastal dune forest, in leaf-litter/soil, leg. J. P. Marais, July 2001 (T 2113/V 9320, one specimen); Lake Sibaya , 27 u 449S, 32 u 239E, coastal bush on dune ridge between lake and sea, in leaf-litter/soil, leg. J. P. Marais, July 2001 (T 2114/V 9285, two specimens); Cape Vidal, Bhangazi Hill, 28 u 07.89S, 32 u 32.89E, 60–80 m a.s.l., dune forest, in leaf-litter, leg. Herbert, Seddon and Tattersfield, Site 2, 27 November 1998 (T 2115/V 7930, 18 specimens); Cape Vidal, Bhangazi Hill, 28 u 07.49S, 32 u 33.39E, 60–80 m a.s.l., dune forest, in leaf-litter, leg. Herbert, Seddon and Tattersfield, Site 1, 27 November 1998 (T2117/W3650, 14 specimens); Cape Vidal, 28 u 07.79S, 32 u 339E, 10–100 m a.s.l., dune forest, in leaf-litter, leg. D. Herbert, 18 October 1997 (T 2036/V 5364, 41 specimens; BMNH 20050568 , two specimens; NMW.Z.2005.035.00002, two specimens; RMNH 101063 About RMNH , two specimens); Greater St Lucia Wetland Park , False Bay Park , 27.96460 u S, 32.37869 u E, closed woodland/forest on fossiliferous sediments beside lake, in leaf-litter and under logs, leg. Earthwatch Team 8, 5 November 2004 (T2049/W2609, three specimens); Hluhluwe Game Reserve, 28 u 04.629S, 32 u 02.79E, 460 m a.s.l., scarp forest, in leaf-litter, leg. Herbert, Seddon and Tattersfield, 29 November 1998 (T 2060/V 7681, two specimens) GoogleMaps .
Other material. Kosi Bay area , near mouth, 26 u 52.1539S, 32 u 52.9559E, dune forest, in leaflitter, leg. S. Lovell et al., 21 October 2003 (W1874) GoogleMaps ; Lake Sibaya , 27 u 449S, 32 u 239E, dune ridge between sea and lake shore, coastal dune forest, in leaf-litter/soil, leg. J. P. Marais, July 2001 (V9273) ; ‘‘ St Lucia Bay’ ’, leg. H. J. Puzey, July 1944 (W1570) ; Mapelane , coastal forest, in leaf-litter, leg. D. Plisko, 14 May 1997 (V5037, V5036) ; Hluhluwe Game Reserve , 28 u 04.59S, 32 u 03.39E, 400 m a.s.l., scarp forest, in leaf-litter, leg. Herbert, Seddon and Tattersfield, 30 November 1998 GoogleMaps (V7660).
Remarks
Specimens of this species have previously been recorded as G. bushmanensis Burnup, 1926 ( Bruggen and Appleton 1977). The material concerned was collected by H. J. Puzey in the St Lucia area, in 1944, and Prof. C. Appleton in the Lake Sibaya area in 1973. Examination of Puzey’s specimens, after return to the Natal Museum from long-term loan, and Appleton’s material housed in the RMNH, showed these to be referable to the present species. This eliminates the record of G. bushmanensis , an otherwise inland mid-altitude species, from coastal localities, a distribution noted as unusual, though not without precedent, by Bruggen and Appleton (1977).
In terms of its apertural dentition and sculpture, G. lindae falls within Group 3B of Herbert and Kilburn (2004). It is, however, very much smaller than any of the other species within this group. Its apertural dentition also resembles that of G. bushmanensis , but that species is much more elongate and almost completely smooth, and its labral tooth is in the form of one bicuspid tooth rather than two separate teeth on a shared base (although the distinctness of these is admittedly somewhat variable in G. lindae ). Gulella maritzburgensis (Melvill and Ponsonby, 1893) , from the KwaZulu-Natal Midlands, is likewise aperturally similar and also has subsutural sculpture, but it is larger (length up to 4.5 mm), has a much more weakly indented suture and its subsutural riblets are extremely fine. Perhaps the most unusual feature of G. lindae is the brevity and relative strength of the subsutural ribbing which gives the suture a milled appearance. This, together with its minute size and five-fold apertural dentition, render the species distinctive.
The distribution of G. lindae resembles that of G. appletoni Bruggen, 1975 , G. browni Bruggen, 1969 , and G. daedalea (Melvill and Ponsonby, 1903) , and it is clearly a tropical element of the fauna. Although currently not recorded from Mozambique, it will undoubtedly prove to occur at least in the southern parts of that country. The northernmost record available (Kosi Bay) lies immediately to the south of the Mozambique border. Whether G. lindae represents an element endemic to the southern extremity of the Indian Ocean coastal belt or is more widespread remains to be established. It was not found in samples collected recently in the Ponta da Barra Falsa area, between Inhambane and Vilanculos, but in general the molluscan fauna of the coastal forests of Mozambique remains almost unknown.
Conservation
Gulella lindae is evidently not rare. Although its currently known area of occupancy is relatively restricted, it is has been found in some numbers in coastal dune forest. Much of this habitat in Zululand falls within the Greater St Lucia Wetland Park World Heritage Site and is thus afforded a high level of protection. Records from inland localities also fall within a conservation area, namely the Hluhluwe-Imfolosi Park.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |