
Turbanella lobata, Yamauchi & Kajihara, 2018
|
publication ID |
https://doi.org/ 10.12782/specdiv.23.183 |
|
publication LSID |
lsid:zoobank.org:pub:3FA0A429-1676-4326-B8D3-FFBA8FE403C9 |
|
persistent identifier |
https://treatment.plazi.org/id/7F0E8CBF-4789-483A-9576-1D65BB4CA1B9 |
|
taxon LSID |
lsid:zoobank.org:act:7F0E8CBF-4789-483A-9576-1D65BB4CA1B9 |
|
treatment provided by |
Felipe |
|
scientific name |
Turbanella lobata |
| status |
sp. nov. |
Turbanella lobata View in CoL sp. nov.
( Figs 7 View Fig , 8 View Fig )
Material examined. Holotype, ICHUM 4982 View Materials (adult), Ishikari beach, Hokkaido, Japan (43°15.420′N, 141°21.438′E), medium-grained sand, 58 cm depth, 5 m landward from high-water level, 14 May 2014 GoogleMaps . Paratype: ICHUM 4983 (subadult), same collection data as for holotype.
Etymology. The specific name is an adjective from the Latin lobatus (lobed), referring to the lateral lobes at U11.
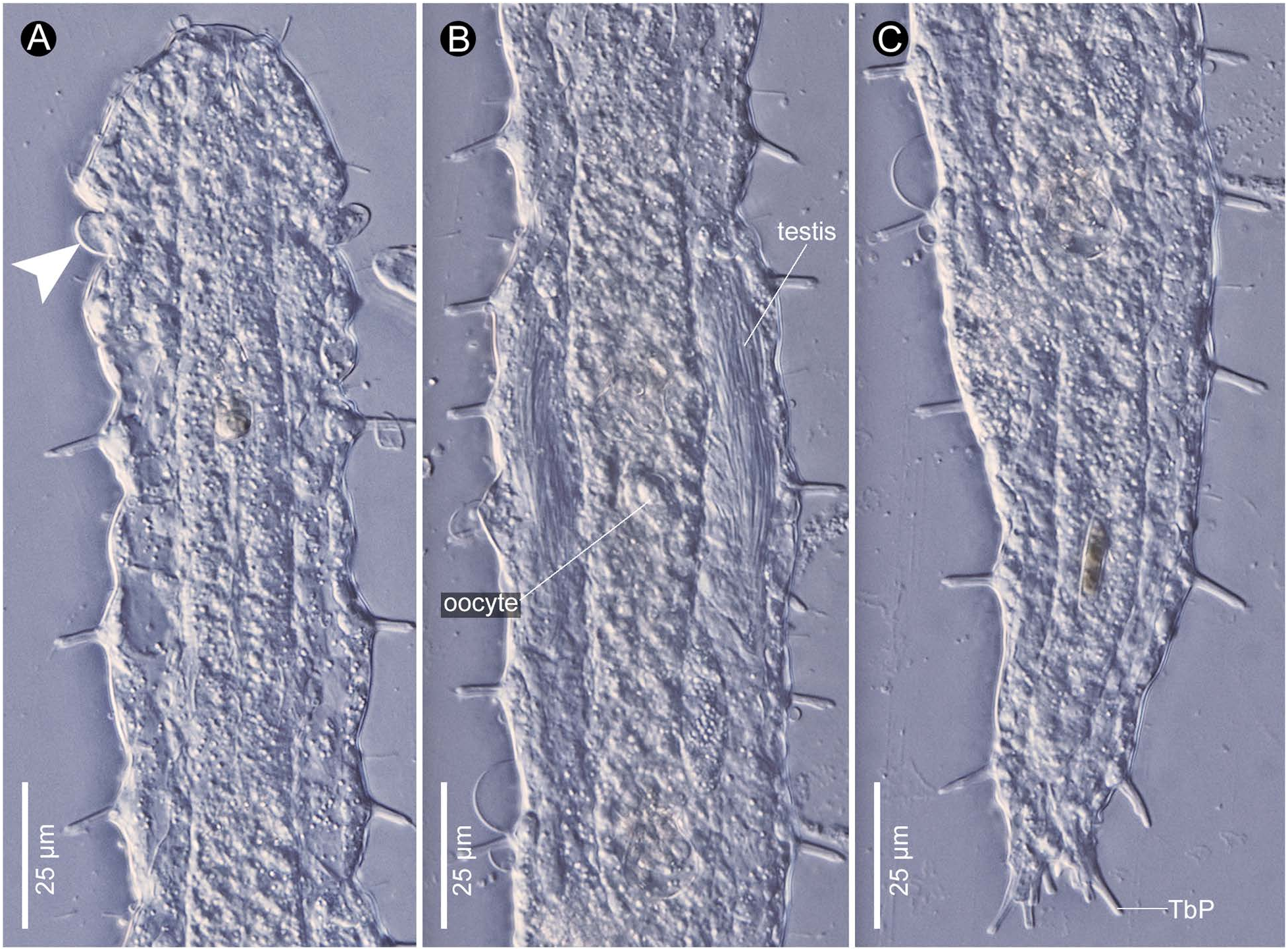
Description. Habitus. Adult Lt 390 µm; L of anterior end to PhJIn (at U30) 132 µm. Body short; head slightly sculpted, with lateral cones at U07 and additional lateral lobes at U11 ( Figs 7A View Fig , 8A View Fig ); neck constriction at U11; trunk widest in mid-body region, tapering gradually to caudal base; caudum moderately cleft, incised from its tips to U97, medial cone absent. Glands 35–40 per side, medium in size (6 µm in diameter), scattered in lateral and medial columns.
Adhesive tubes. TbA four per side (L 5–7 µm), occurring on lobe inserted at U11 ( Fig. 7B View Fig ); TbL 10–12 per side (L 8–12 µm), some bearing cilia, irregularly spaced and often asymmetrically arranged, with tow in pharyngeal region, one behind anal opening, and others along intestine; TbD 7–8 per side, with one in pharyngeal region and remainder along intestine; ‘cirrata’ [Seitenfüsschen] tubes occurring at U38; TbP four per side, arrayed along rear edge of each lobe, lengthening medial to lateral (L 3–9 µm) ( Figs 7A, B View Fig , 8C View Fig ).
Ciliation. Mouth surrounded by short sensory cilia (L 3 µm), with longer cilia (L 5 µm) inserted at points of head sculpting on each side; ciliary hairs (L 11 µm) forming cir- cum-cephalic band at U07; sensory cilia (L 9 µm) occurring on trunk in lateral columns; each Tb inserted on trunk, bearing cilium (L 11 µm) arising from rear apex of tube support; ventral locomotory cilia (L 12 µm) running in two longitudinal bands along lateral body margins to anus ( Fig. 7B View Fig )
Digestive tract. Mouth terminal, of medium width (11 µm); buccal cavity conical-shaped; walls lightly cuticular; pharynx of medium width throughout, with basal pharyngeal pores at U30; intestine narrowing anterior to posterior; anus at U91.
Reproductive tract. Hermaphroditic; paired testes extending posteriorly from U59, their vasa deferentia recurving anteriorly, but terminal not seen; bilateral oocytes developing posterior to anterior, largest (53×23 µm) in anterior region of intestine ( Figs 7A View Fig , 8B View Fig ).
Remarks. Among approximately 30 species in the genus Turbanella , five species share many features with Turbanella lobata sp. nov.: T. caledoniensis Hummon, 2008 ; T. lutheri Remane, 1952 ; T. otti Schrom, 1972 ; T. pacifica Schmidt, 1974 ; and T. subterranea Remane, 1934 . These species differ from T. lobata sp. nov. as follows: T. caledoniensis lacks lateral head lobes and the neck constriction; T. lutheri banelloides was originally established in Paradasys ( Boaden 1960) , suggesting that transfer of the species to Cephalodasys ( Hummon 1974) may require a revision. Our analysis indicates Cephalodasys and Cephalodasyidae as currently diagnosed ( Hummon and Todaro 2010; Kieneke et al. 2015) are not monophyletic ( Fig. 4 View Fig ). However, additional gene markers may recover them as monophyletic, because support values for basal nodes are generally low ( Fig. 4 View Fig ). In any case, inclusion of C. maximus Remane, 1926 , the type species of Cephalodasys , as well as P. subterraneu , the type species of Paradasys (see Remarks for C. mahoae sp. nov. above), in molecular phylogenetic context is indispensable to test the appropriateness of the generic placement of C. mahoae sp. nov., as well as for taxonomic revision of the family.


| ICHUM |
Invertebrate Collection of the Hokkaido University Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.