
Tlayuamichin itztli
|
publication ID |
https://doi.org/ 10.5281/zenodo.205966 |
|
DOI |
https://doi.org/10.5281/zenodo.6184215 |
|
persistent identifier |
https://treatment.plazi.org/id/0399C467-FFE3-021E-FF42-FF1C6FA0FA93 |
|
treatment provided by |
Plazi |
|
scientific name |
Tlayuamichin itztli |
| status |
|
Tlayuamichin itztli gen. et sp. nov.
( Figs 2–15 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 View FIGURE 11 View FIGURE 12 View FIGURE 13 View FIGURE 14 View FIGURE 15 )
Lepidotes sp. Applegate, 1996: 531, fig. 1
Holotype. IGM 6716; fairly complete specimen of 403 mm SL ( Fig. 2 View FIGURE 2 ). Although missing parts of the unpaired fins, the specimen is almost completely and excellently preserved.
Paratypes. IGM 6717–5 ( Figs 3–5 View FIGURE 3 View FIGURE 4 View FIGURE 5 ). IGM 6717 is an almost complete specimen, but preserved in lateroventral view and, although the bones are not largely displaced, the skull is disarticulated ( Fig. 3 View FIGURE 3 A). IGM 6718 is missing the head and pectoral girdle, but the dorsal, anal, and caudal fins are very well preserved ( Fig. 3 View FIGURE 3 B). IGM 6719 contains a very well preserved and articulated skull with the anterodorsal portion of the body without fins ( Fig. 4 View FIGURE 4 ). IGM 6720 is partially incomplete (the paired fins and part of the unpaired fins and squamation are missing) but, although some of the bones of the head are crushed, its general preservation is excellent displaying some important skull features not preserved in the holotype ( Fig. 5 View FIGURE 5 ).
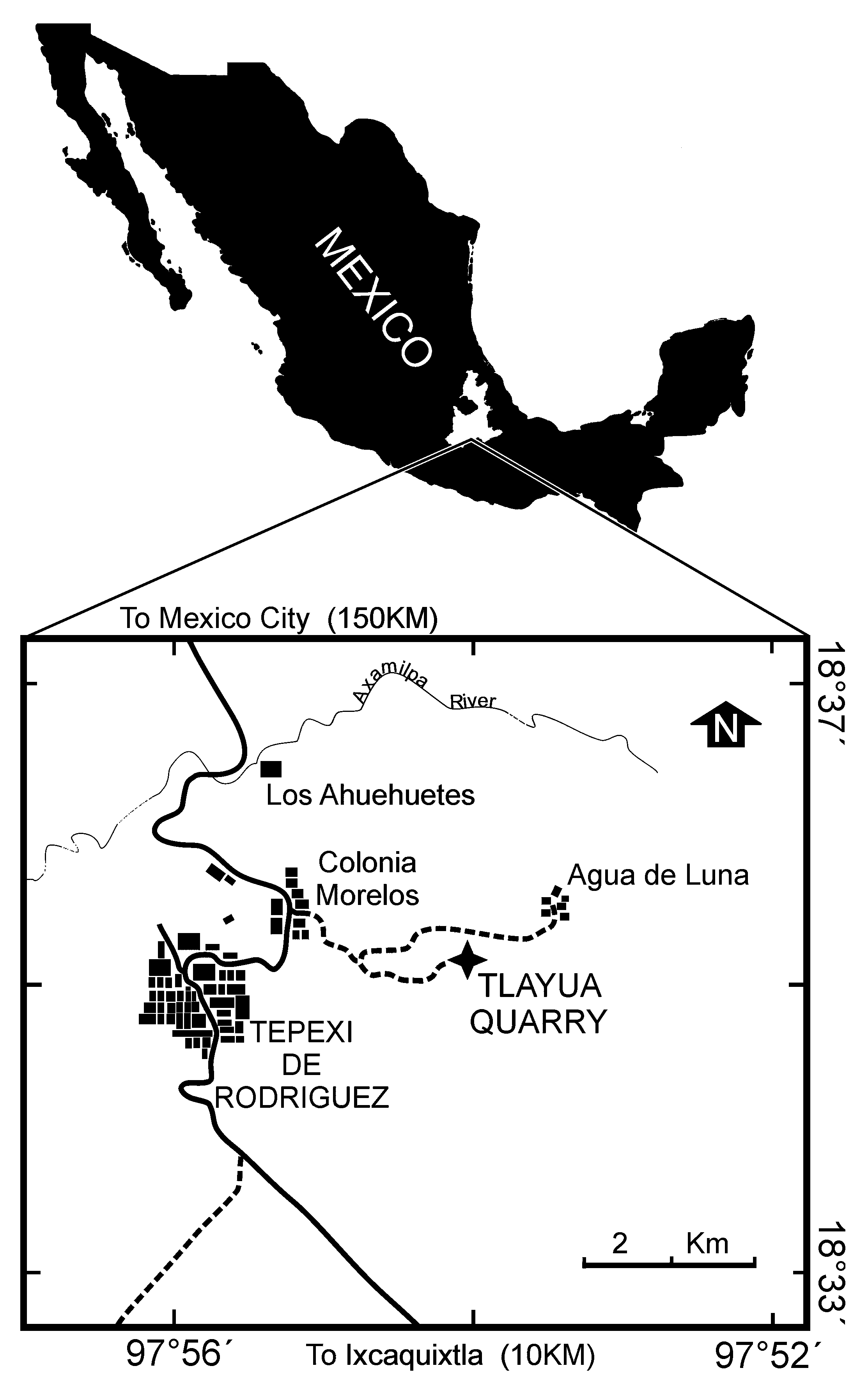
Type locality. Tlayúa quarry, near Tepexi de Rodríguez, Puebla, Mexico ( Fig. 1 View FIGURE 1 ).
Type horizon. Middle Member of the Tlayúa Formation ( Pantoja-Alor, 1992); Middle-Upper Albian (Early Cretaceous) ( Seibertz & Buitrón 1987; Cantú-Chapa 1987; Seibertz & Spaeth 2002; Benammi et al. 2006).
Derivation of name. The name of the fish is also written in the Náhuatl language. The species epithet “ itztli ” means “obsidian” and it refers to the scale color in this fish. The complete generic and species name of this semionotid means “the fish of Tlayúa with obsidian color scales”.
Diagnosis. Frontals very long, about 4,5 times longer than wide, and narrowing anteriorly up to 37% of their maximal width; antorbital process absent; dermopterotic with anteroventral process articulating with the sphenotic*; operculum does not contact the dermopterotic; a series of paraethmoids anterior to the supraorbital series*; supraorbital series extends beyond the anterior rim of the orbit*; large pentagonal supraorbital partially closing the anterior rim of the orbit*; seven anterior infraorbitals*; infraorbitals at the ventral border of the orbit expanded dorsally, acuminating anterodorsally*; dorsalmost suborbital separating the preoperculum from the dermopterotic; premaxillary teeth smaller than other jaw or palatal teeth*; modified pectoral scale present.
Pterygial formula: D23 T34
P7 A18 C28
Description. Tlayuamichin itztli is a fusiform fish with gently curved dorsal and ventral borders. Although the SL in IGM CT 1 and IGM CT 2, the most complete specimens available, is 42.2 and 27.6 cm respectively; Tlayuamichin itztli could be larger as shown by IGM CT 2, in which the SL could be around 44.6 cm. The maximal depth of the body (MBD), measured midway between the insertion of the pectoral and pelvic fins, is 0.39–0.4 of the SL. The head is triangular and slightly deeper than long. The maximal depth of the head (MDH) enters about 3.6 times in the SL and is almost 1.7–1.9 times the depth of the caudal peduncle. The length of the head (HL) is 0.3 of the SL and 5–5.5 times the longitudinal diameter of the orbit. The paired fins are sub-abdominal, the pelvic fin being located just behind the middle of the body, at 0.52 of the SL. Both unpaired fins are short and located in the posterior half of the body. The dorsal fin inserts at the 24rd raw of scales, at 0.6 of the SL, and the anal fin at the 17th raw of scales, at 0.68–0.74 of the SL. Body proportions are based on the specimens IGM 6716, 6718 and 6720.
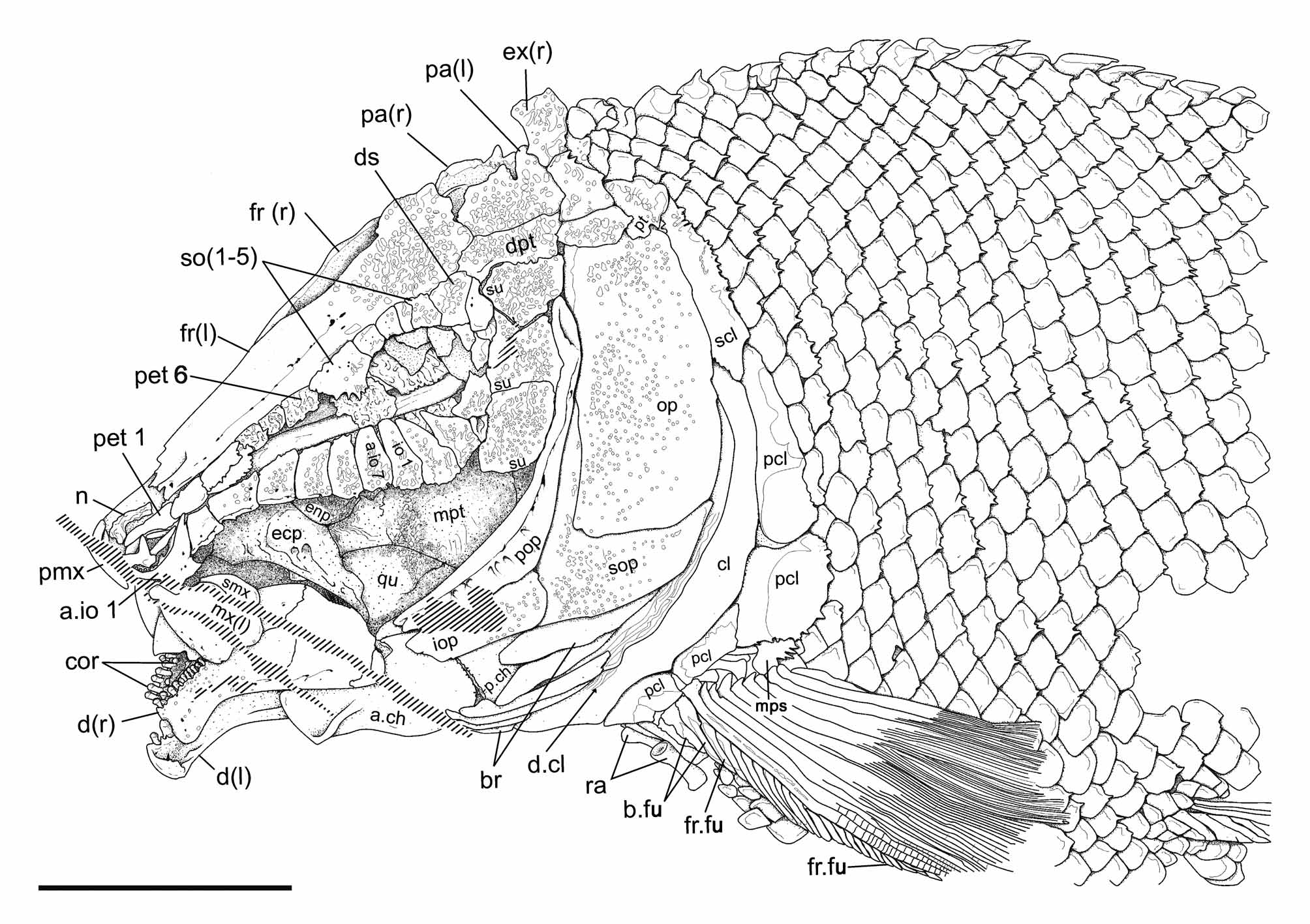
Skull roof. The bones of the skull roof are densely ornamented with tubercles. The ornamentation is more dense in the posterior part of the skull, on the extrascapulars, parietals, dermopterotics and posterior part of the frontals. Several tubercles in this area preserve a very thin layer of ganoin. There are two (left side of IGM 6716; Figs 6–7 View FIGURE 6 View FIGURE 7 ) or three (right side of IGM 6719 and 6720; Figs 4 View FIGURE 4 , 8–9 View FIGURE 8 View FIGURE 9 ) extrascapular bones on each side of the skull. When two extrascapulars are present, the medial extrascapulars are larger than the lateral ones, and are mediolaterally elongate, narrower medially than laterally. When three extrascapulars are present, the two most medial elements occupy an area equivalent to the single medial extrascapular described before. The lateral extrascapulars are trapezoidal, a little narrower dorsally than ventrally. The parietals are rectangular, twice as long as wide, and about a third of the length of the frontals. They form a sinuous inter-parietal suture. In the holotype, the inter-parietal suture has an indentation to the left, but in IGM 6719 there is no indentation in this suture ( Fig. 4 View FIGURE 4 ). The parietals articulate with the frontals anteriorly, laterally with the dermopterotic, and posteriorly they meet the median extrascapulars only. In IGM 6719 the lateralmost extrascapular apparently articulates with the parietal, but this is due to the post-mortem ventral displacement of the parietal and dermopterotic ( Fig. 9 View FIGURE 9 ). The frontals are very long, about 4,5 times longer than wide and articulate with each other through a sinuous suture. The frontals are widest at the posterodorsal corner of the orbit and the posterior border is rounded. Anteriorly, the frontal narrows gradually up to 37% of it maximal width and there is no antorbital process. The nasal bones are well preserved in the holotype (IGM 6716; Figs 6–7 View FIGURE 6 View FIGURE 7 , 10 View FIGURE 10 ) and in IGM 6719. They are delicate, longitudinally elongate bones with laterally curved anterior ends.
The exposed portion of the dermopterotic is rectangular, about 3 times longer than wide. This bone articulates with the frontal anterodorsally, with the lateral extrascapular posteriorly, with the parietal dorsally, and with a large suborbital anteroventrally. In Tlayuamichin itztli the dermopterotic does not contact the anterodorsal corner of the operculum, as it is the case in many other semionotiforms. The anterior portion of the ventral margin of the dermopterotic is distinctly concave and the bone is expanded in an anteroventral process that articulates medially with the sphenotic and is superficially overlapped by the dermosphenotic ( Fig. 9 View FIGURE 9 A–B).
Braincase and parasphenoid. Although no isolated braincase is preserved, some braincase structures are exposed in the holotype mainly. The sphenotic forms a strong lateral process that is projected laterally and presents a dermal component, which is superficially exposed between the dermosphenotic, the last infraorbital bone, and the suborbital bones ( Figs 6–7 View FIGURE 6 View FIGURE 7 ). This small tear-shape dermal component of the sphenotic is ornamented with tubercles covered with a very thin layer of ganoin.
The ethmoidal and orbital regions of the braincase are partially preserved. Tlayuamichin itztli peculiarly has a series of apparently endochondral bones aligned lateral to the very elongate ethmoidal region and above the parasphenoid. The series of paired bones extend from the first supraorbital to the tip of the snout dorsal to the antorbital, and these bones, six in total, show a serial correspondence with the anterior infraorbital bones ( Figs 6–7 View FIGURE 6 View FIGURE 7 , 10 View FIGURE 10 ). The endochondral nature of these bones is indicated by the state of preservation, but it is not possible to certify this condition. Also, their actual origin is unknown and very difficult to interpret due to the lack of information. Such bones are so far unknown in other actinopterygians and, thus, comparisons are not possible. They might be ossifications originated from the ethmoidal cartilage, probably homologous with the lateral ethmoids. Since these bones are placed laterally along the elongate ethmoidal region, we propose to name them paraethmoids, indicating their topographic relationships. The paraethmoids are well preserved in the holotype (IGM 6716) only, where the paraethmoids 3 and 4 are very damaged due to the intrusion of two displaced anterior infraorbitals from the right side of the skull. In IGM 6720 ( Fig. 8 View FIGURE 8 ) the most posterior of these elements is probably laterally covered by the most anterior dermal supraorbital (this supraorbital is missing in the holotype).
Some partially preserved chondral elements within the orbit most probably represent remains of the orbitosphenoid. The orbital part of the parasphenoid is exposed in the holotype and in IGM 6719 and 6720 ( Figs 4 View FIGURE 4 , 8 View FIGURE 8 ). In IGM 6717 the parasphenoid is completely exposed, though poorly preserved in ventral view. The parasphenoid is edentulous and extends back throughout the area corresponding to the prootics and ends at the central portion of the basioccipital forming two posterior processes. There is a longitudinal deep median groove along the dorsal surface of the orbital section of the parasphenoid. Within the orbital region, the parasphenoid broadens posteriorly, reaching its maximal breadth at the posterior wall of the orbit, where there are two long and narrow basipterygoid processes. Dermal and possible chondral portions of the basipterygoid processes cannot be distinguished, but the processes are very large.
Circumborbital series. The circumorbital series, which is completely preserved in the holotype (IGM 6716; Figs 6–7 View FIGURE 6 View FIGURE 7 ) and IGM 6720 ( Fig. 8 View FIGURE 8 ), involves five supraorbitals, the dermosphenotic, six infraorbitals (io1––io 6 in anterior to posterior direction), seven anterior infraorbitals (a.io1–a.io 7 in anterior to posterior direction) and the antorbital. The circumborbital series is open (IGM 6716 and 6719; Figs 4 View FIGURE 4 , 6–7 View FIGURE 6 View FIGURE 7 , 10 View FIGURE 10 ), though it appears to be closed in IGM 6720 ( Fig. 8 View FIGURE 8 ) due to the anterodorsal displacement of the bones in the anteroventral portion of the skull. Tlayuamichin itztli is very peculiar in having the most anterior supraorbital element very large, placed exactly at the anterior rim of the orbit so that the bone, which has the shape of an inverted pentagon, has equally large orbital and anteorbital portions (it should be noticed that in IGM 6716 the bone is fractured right at the level of the anterior rim of the orbit, giving the appearance of two separate ossifications; Figs 6–7 View FIGURE 6 View FIGURE 7 , 10 View FIGURE 10 ). This pentagonal supraorbital forms a ventral wedge that faces a dorsal wedge formed by the last anterior infraorbital (a.io7) and the first infraorbital (io1) together, so that the two opposite wedges are partially closing the orbit anteriorly. In IGM 6720 there is another additional supraorbital anterior to this pentagonal bone ( Fig. 8 View FIGURE 8 ). There are no traces of such an additional supraorbital in the other specimens, but it might have been present and not preserved. In any case, Tlayuamichin itztli is very peculiar in having a long series of supraorbital bones, which extends beyond the orbit anteriorly. The remaining supraorbitals are rectangular and relatively small bones. The supraorbital bones are ornamented with tubercles, which are covered with a very thin layer of ganoin.
As usual in semionotiforms, the dermosphenotic is located at the posterodorsal corner of the orbit. This bone is rectangular, about twice as deep as long, and as long as the adjacent supraorbital ( Figs 4 View FIGURE 4 , 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 ). The dermosphenotic is also ornamented with tubercles covered with a very thin layer of ganoin.
The infraorbital bones are best preserved in the holotype (IGM 6716; Figs 6–7 View FIGURE 6 View FIGURE 7 ). Among the six infraorbitals, three form the ventral margin of the orbit (infraorbitals io1–io3), the fourth infraorbital (io4) is placed at the posteroventral corner of the orbit and the last two infraorbitals (io5–io6) form the posterior margin of the orbit. The infraorbitals three to six (io3–io6) articulate with suborbital bones posteriorly. The infraorbitals five and six (io5– io6) are relatively small, deeper than long, rectangular bones. The infraorbital four (io4) is about twice the size of the infraorbital five (io5) and smaller than the infraorbital three (io3); it has a concave orbital border and is convex posteriorly. The infraorbitals one to three (io1–io3) are irregularly triangular, acuminated anterodorsally. In the holotype, the infraorbital three (io3) is slightly rotated clockwise and displaced dorsally because of the extrusion of the basipterygoid process of the parasphenoid. When moved back to its natural position, the infraorbital three (io3) alone forms most of the ventral margin of the orbit. The same happens in IGM 6720. The infraorbital two (io2) is the deepest bone of the whole series, followed by the infraorbital one (io1), which is only slightly shallower, and the infraorbital three (io3), which is about 20% shallower than the infraorbital two. Accordingly, the ventral margins of the infraorbitals 1–3 together describe a gently convex curve.
The seven anterior infraorbitals are aligned ventral to the level of the parasphenoid ( Figs 6–7 View FIGURE 6 View FIGURE 7 , 10 View FIGURE 10 ). The anterior infraorbitals four to seven (a.io4–a.io7) are approximately rectangular, deeper than long, with their depths gradually decreasing anteriorly. The anterior infraorbital 7 is slightly acuminated dorsally, in the anterior infraorbital 6 the dorsal border is only slightly shorter than the ventral border, and the dorsal and ventral borders of the infraorbital 5 are approximately equal. The anterior infraorbital 4 is more irregular in shape, with the posterior border deeper than the anterior border and has, thus, anteroventrally inclined dorsal border. The anterior infraorbital 3 is approximately rectangular, about 1,5 times longer than deep, with the dorsal border a little longer than the ventral border. The anterior infraorbital 2 is smaller than its adjacent elements in the series. It is slightly narrower dorsally than ventrally and has a small laminar outgrowth ventral to the passage of the infraorbital sensory canal. Such an outgrowth is not present in the more posterior anterior infraorbitals. The first anterior infraorbital is somewhat boomerang-shaped and also has a well-ossified outgrowth ventral to the passage of the infraorbital sensory canal.
All the infraorbital and anterior infraorbital bones are also ornamented with tubercles covered with a very thin layer of ganoin, which are denser towards the ventral and posteroventral border of the circumborbital ring.
The most anterior bone of the infraorbital series is usually interpreted as the antorbital. In Tlayuamichin itztli this most anterior element resembles the small, almost tubular antorbital in “ Lepidotes ” minor illustrated by Patterson (1975: fig. 135) ( Figs 6–7 View FIGURE 6 View FIGURE 7 ). An irregularly shaped ossification preserved anterior to the antorbital and dorsal to the anterior end of the first anterior infraorbital might represent a displaced rostral.
Suborbital bones. There are three suborbital bones in Tlayuamichin itztli , and they are also densely ornamented with tubercles covered with a thin layer of ganoin. They are all of similar size and aligned in a single row completely covering the area between the circumborbital bones and the preoperculum, but do not extend anteriorly, ventral to the orbit. The most dorsal suborbital is expanded posteriorly and completely separates the preoperculum from the dermopterotic ( Figs 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 ).
Opercular bones. The preoperculum is a crescent-shape bone, very narrow and gently curved, narrowing both dorsally and anteriorly ( Figs 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 ). Anteriorly, the preoperculum extends beyond the anterior margin of the orbit. Dorsally, the preoperculum does not reach the dermopterotic; its dorsal tip being widely separated from that bone by the most dorsal suborbital. The operculum is large and subrectangular, a little narrower dorsally than ventrally. It is twice as deep as long in the holotype and in IGM 6720 ( Figs 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 ), but only 1,6 times deeper than long in IGM 6717 ( Fig. 3 View FIGURE 3 ) and, thus, the shape of this bone is somewhat variable within the species. There are three irregularly shaped opercular ossifications in IGM 6719: a large dorsal operculum, an even larger ventral operculum, and a small rounded ossification at the anteroventral end of the ventral operculum ( Fig. 9 View FIGURE 9 ). The three bones occupy together an area about 1,5 times deeper than long, but the dorsal operculum is about 0,6 times narrower than the ventral operculum. The suboperculum has a very high ascending process, which is about 70% of the maximal length of the bone (completely exposed only in the holotype; Figs 6–7 View FIGURE 6 View FIGURE 7 ). The ascending process forms an interior angle of about 90° with the dorsal border of the suboperculum. In IGM 6716, 6717 and 6720 the suboperculum is deepest anteriorly, where it reaches about 30% of the depth of the operculum, and tapers posteriorly with curved dorsal and ventral borders ( Figs 3 View FIGURE 3 , 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 ). In IGM 6719 the suboperculum is almost equally deep throughout its length and forms a sinuous articulation with the ventral operculum ( Fig. 9 View FIGURE 9 ). The ascending process of the suboperculum in this fish is hidden by the somewhat displaced preoperculum. Anteriorly, the suboperculum articulates with the interoperculum through an almost vertical suture. The interoperculum extends anteriorly up to the level of the anterior end of the preoperculum, thus extending medioventral to the whole ventral margin of that bone. The ventral border of the interoperculum is gently convex. All the opercular bones are ornamented with numerous tubercles covered with ganoin, except for the preoperculum where the tubercles are only very few and scattered. It is worthy to note that the peculiarities of the opercular bones in IGM 6719 described above are considered as abnormalities. IGM 6719 presents multiple ossification of the operculum and probably also de preoperculum and an abnormal shape of the suboperculum. Although it is important to report such cases, in the lack of ontogenetic information it is impossible to know the causes or processes that might led to such deviations in this fish.
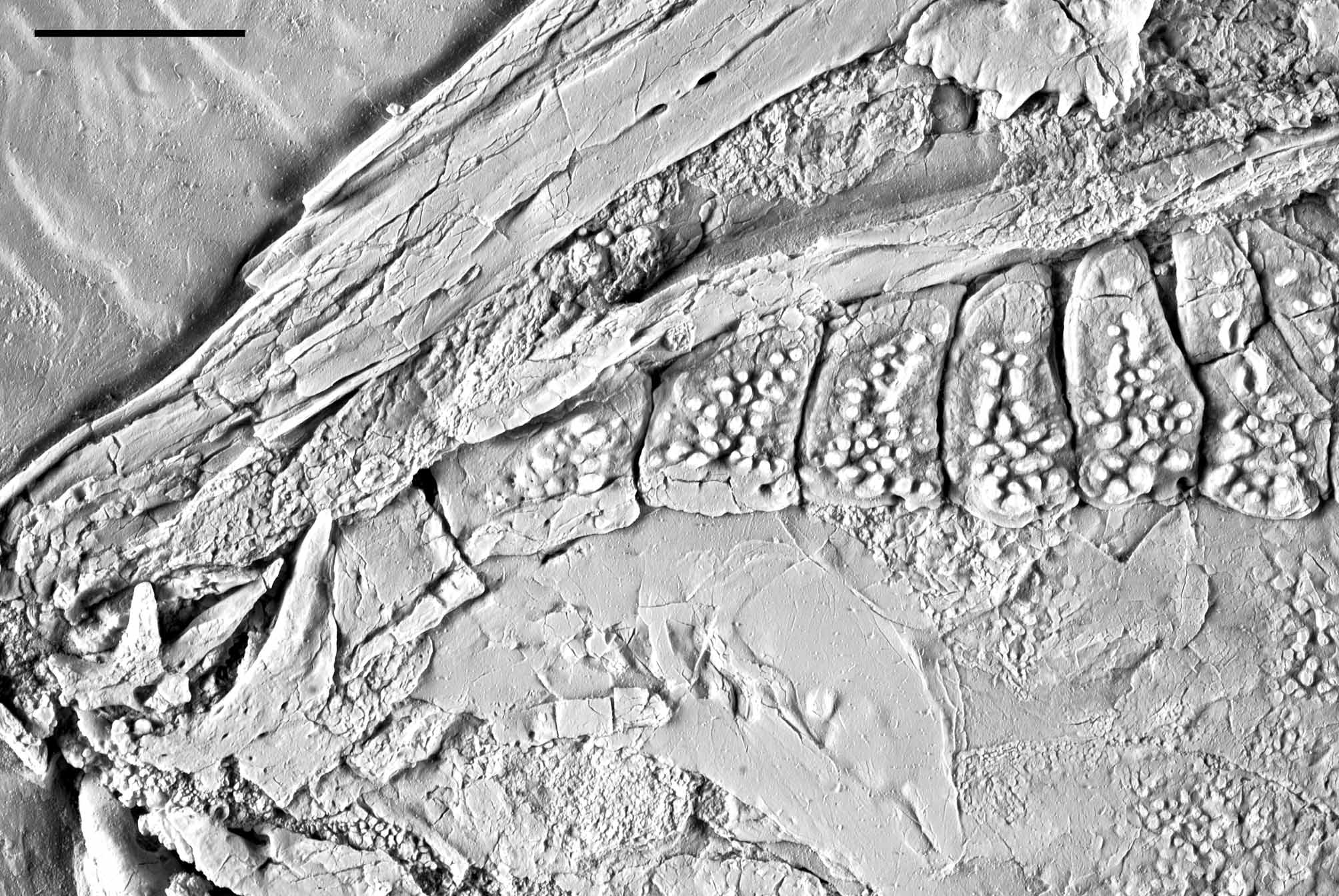
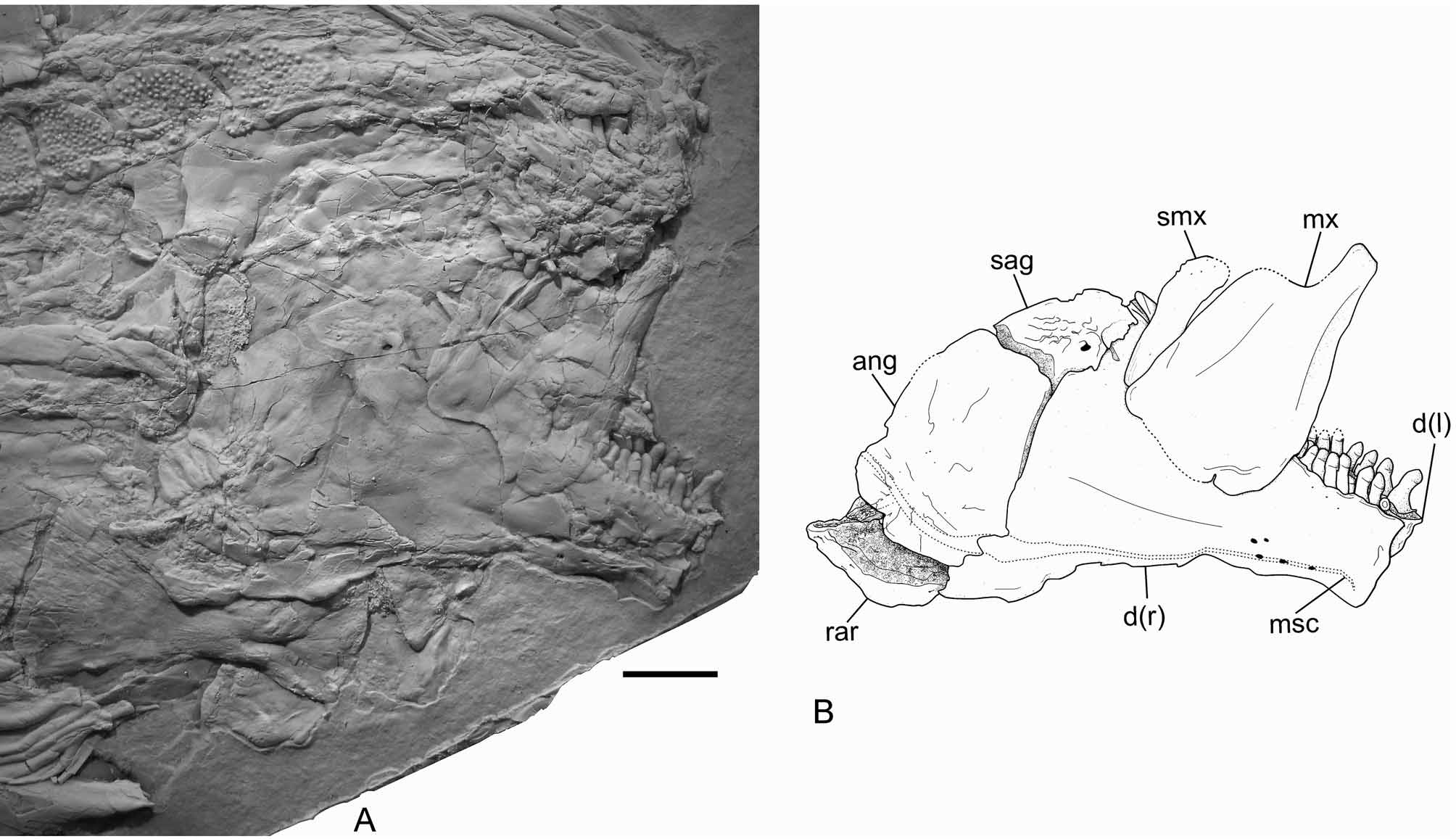
Jaws. The jaws are best preserved in IGM 6719 ( Fig. 11 View FIGURE 11 ), but they are also relatively well preserved in the holotype and IGM 6717 ( Figs 3 View FIGURE 3 , 6–7 View FIGURE 6 View FIGURE 7 , 12 View FIGURE 12 ). The lower jaw joints the quadrate at the level of the anterior orbital margin. It is a triangular structure, about 2.5 times longer than deep. The posterior and dorsal borders of the lower jaw are convex, the ventral border bents ventrally at the alveolar portion, and the alveolar border is straight, inclined anteriorly. The dentary symphysis is straight, vertical, and is contained about 2.5 times in the maximum depth of the coronoid process and more than 5 times in the total length of the lower jaw. The lower jaw is only exposed in lateral view in the available specimens, and the dentary, angular, surangular and retroarticular bones are recognized. The dentary is large and elongate, with a short posteroventral process, which is only about 15 % of the total length of the lower jaw. The posterior border of the dentary, which articulates with the angular, forms an obtuse angle with the posteroventral process. Small tubercles ornament the external lateral surface of the anterior portion of the dentary below the alveolar border. There is a single row of dentary teeth, which have short pedicles and rounded crowns. Of the right side of the skull, two coronoid bones, anterior and posterior, are exposed in IGM 6716 ( Figs 6–7 View FIGURE 6 View FIGURE 7 ). Only the posterior of these coronoid bones is exposed on the left side of this specimen. The coronoids bear only one raw of teeth, which are identical to the marginal teeth of the dentary. The angular articulates with the dentary between the coronoid process and the posteroventral process, with the retroarticular posteroventrally, and with the surangular anterodorsally ( Fig. 11 View FIGURE 11 ). The angular is excavated for the articulation of the quadrate posteriorly. This articular facet is concave and well distinct, but not deeply excavated. As usual in semionotiforms, the surangular places on top of the lower jaw articulating dorsal to the angular and dentary, and it is dorsally convex. The retroarticular is relatively large and well ossified, forming the posteroventral end of the lower jaw. It articulates with the posteroventral process of the dentary anteriorly, and with the angular anterodorsally.
The upper jaw is also shown in IGM 6716, 6717 and 6719 ( Figs 6–7 View FIGURE 6 View FIGURE 7 , 11 View FIGURE 11 , 12 View FIGURE 12 ). The premaxillae are badly preserved; in IGM 6716, only the nasal processes of both premaxillae are exposed. In IGM 6717 the two premaxillae are preserved in ventral view ( Fig. 12 View FIGURE 12 ). The alveolar portion of the premaxilla is narrow. The exact number of premaxillary teeth is unknown; three alveoli are preserved in the right premaxilla and four in the left premaxilla, but in both cases the bone is broken and more alveoli might have been present. According to the size of the alveoli, the premaxillary teeth are notably smaller than the vomerine or dermopalatine teeth and even fairly smaller than the dentary teeth. Therefore, the premaxillary teeth are the smallest in the jaws and palate. The maxilla is best preserved in IGM 6719 and is an edentulous flat bone. The articular process is strong, rod-like and long (broken in IGM 6719, Fig. 11 View FIGURE 11 , but complete in IGM 6717, Fig. 12 View FIGURE 12 ). Immediately posterior to this process, the maxilla expands abruptly dorsally forming a subrectangular plate with rounded corners, which is about 1.2 times longer than deep ( Fig. 11 View FIGURE 11 ). The posterior border of the maxilla is almost well preserved in IGM 6719 only. However, the border is broken and, thus, it was reconstructed in Figure 11 View FIGURE 11 B according to the imprint left on the dentary. The posterior bor- der of the maxilla is therefore almost straight and immediately below the broken piece there is a sharp indentation close to the posteroventral corner of the bone, which is not due to any kind of fracture or brake. A long and slender supramaxilla lies along the whole dorsal border of the maxillar plate.
Palatoquadrate ossifications. In Tlayuamichin itztli the cheek is naked and the palatoquadrate ossifications are best exposed and well preserved in the holotype (IGM 6716), which shows large part of the quadrate, ectopterygoid, metapterygoid, and only some parts of the entopterygoid ( Figs 6–7 View FIGURE 6 View FIGURE 7 ). The quadrate is fan shaped with convex dorsal border and the articular process directed ventrally. The articular process of the quadrate is small and short. Only the most anterior portion of the quadratojugal is visible in IGM 6716 attached to the ventral border of the quadrate and far back far from the articular facet for the lower jaw. The ectopterygoid articulates with the whole anterior border of the quadrate; its shape is crescent, with gently convex dorsal and concave ventral borders, and it is about two times longer than the quadrate. The anterior portion of the ectopterygoid is broken in the holotype and no teeth are preserved in this area. The two dermopalatines are well exposed in IGM 6717 ( Fig. 12 View FIGURE 12 ). The bones are broad, rounded, and bear 7–9 teeth irregularly ordered. IGM 6717 also shows the paired vomers, which are still articulated. Each vomer bears 4–5 irregularly arranged teeth. These palatine and vomerine teeth are similar to those on the jaws, also with relatively short pedicles and rounded crowns, though somewhat larger than them. The entopterygoid is also exposed in IGM 6717 and it is completely edentulous. Only the posterior portion of the entopterygoid is exposed in IGM 6716 suturing with the ectopterygoid and metapterygoid, but not with the quadrate ( Figs 6– 7 View FIGURE 6 View FIGURE 7 ). The metapterygoid is very broad, meeting the dorsal border of the entopterygoid anteriorly and the ectopterygoid and quadrate ventrally ( Figs 6–7 View FIGURE 6 View FIGURE 7 ).
Hyoid arch and branchiostegal rays. The hyoid arch is partially shown in IGM 6716, 6717, and 6719 ( Figs 3 View FIGURE 3 A, 4, 6–7, 12). The hyomandibula is not shown in any of the available specimens. The anterior ceratohyal is relatively large and slightly waisted. Its posterior border is about two times deeper the anterior one. The constriction or waist divides the anterior ceratohyal in two portions, the posterior portion being twice as long as the anterior portion. The longitudinal axis and the anterior and posterior borders of the anterior ceratohyal are massively ossified, whereas the rest of the bone is laminar. The posterior ceratohyal is partially shown in the specimens mentioned above. The anterior and posterior ceratohyals articulate through cartilage and a superficial medial irregular suture. No hypohyal or basihyal is preserved, not even in IGM 6717, which preserves the skull in ventral view and, thus, these bones probably remain cartilaginous.
The branchiostegal rays are preserved partially articulated in IGM 6716 and 6719 ( Figs 4 View FIGURE 4 , 6–7 View FIGURE 6 View FIGURE 7 ). Considering the two specimens, the following arrangement is interpreted: five slender branchiostegal rays articulate with the anterior ceratohyal, and two larger, plate-like branchiostegals articulate with the posterior ceratohyal. The most posterior branchiostegal ray is overlapped by the suboperculum and has approximately the same length as the ventral margin of that bone, and about three times the length of the most anterior and smallest branchiostegal ray.
Sensory canals of the head. The trajectories of the cephalic sensory canals are best preserved in IGM 6716 and 6719 ( Figs 4 View FIGURE 4 , 6–7 View FIGURE 6 View FIGURE 7 ). The supraorbital sensory canal is represented by a groove that is perforated from time to time, throughout the central portion of the nasals, the lateral portion of the frontals and the central portion of the anterior halves of the parietals. In the parietals, the anterior, middle and posterior pit lines are also represented by perforated grooves.
The temporal sensory canal is represented by some relatively large pores on the anterior and ventral margins of the dermopterotic, and on the lateral margin of the most lateral extrascapular. In the extrascapulars, the supraoccipital commissure is also represented by pores, but it is not possible to know whether it was continuous from one to the other side of the skull.
The supraorbital bones are all pierced by foramina that are interpreted as the orbital sensory canal as described in López-Arbarello and Sferco (in press). The orbital canal meets the temporal and infraorbital canals in the dermosphenotic. The trajectory of the infraorbital sensory canal is represented by grooves in several infraorbital bones. In the small infraorbital bones placed at the posterior border of the orbit, the sensory canal probably traversed their thickened anterior margin. More anteriorly, the grooves representing the canal are shown in the ventral half of the infraorbital bones. From the posteroventral corner of the orbit forwards, the grooves are gradually closer to the ventral border of these bones. Additionally, a series of relatively large pores at the ventral margin of the infraorbital bones most probably represent the exits of branches of the infraorbital sensory canal. In the antorbital the infraorbital canal is only represented by two pores placed at its ventral margin. As mentioned above, a small bone traversed by a groove and placed anterior to the antorbital probably represents a rostral traversed by the ethmoidal commissure.
The preopercular sensory canal apparently traversed the thickened anterior margin of the preopercular bone. At least 8 pores distributed throughout the central portion of the preoperculum in IGM 6716 indicate the exits of branches from this sensory canal. In the most dorsal suborbital there is a pore at the ventral margin, placed directly opposite to the thickened anterior margin of the preoperculum, and followed by a superficial slight groove that ends opposite to a pore at the ventral margin of the dermopterotic ( Fig. 9 View FIGURE 9 C). These structures indicate that the preopercular sensory canal traversed the posterior portion of the most dorsal suborbital, a bone that, as explained above, completely separates the preoperculum from the dermopterotic.
Anteriorly, the preopercular sensory canal continues as the mandibular sensory canal through the angular and the dentary. In these bones, the mandibular canal is represented by several fenestrae and a groove running close to the ventral border of the lower jaw. Additional pores in one or the other specimen are not constantly present and might represent branches of the main canal or might have a different origin (e.g. innervation or nutrition).
Pectoral girdle. The dermal bones of the pectoral girdle are best preserved in the holotype (IGM 6716; Figs 6– 7 View FIGURE 6 View FIGURE 7 ), in which the left girdle is complete and articulated, including a posttemporal bone, a supracleithrum, a cleithrum, probably a serrated organ and four postcleithra. As usual in semionotiforms the posttemporal has an approximately triangular outline, narrowing dorsally. The bone is densely ornamented with ridges and/or tubercles covered with a thin layer of ganoin, and ends dorsally at the level of the dorsal border of the median extrascapular bone when three extrascapular bones are present. The anteroventral process of the posttemporal is short and broad, laterally compressed. The posteroventral border of the posttemporal is convex and articulates with a concave surface in the supracleithrum.
The supracleithrum is dorsoventrally elongate and the exposed portion is relatively narrow. The supracleithrum is overlapped by the posttemporal dorsally and the operculum anteriorly, and it ventrally overlaps the cleithrum and the most dorsal postcleithrum. The posterior border of the supracleithrum is straight through most of its length, until meeting the most dorsal postcleithrum, where it bents anteroventrally, so that the bone tapers in that direction. Only dorsal to the postcleithrum, the posterior margin of the supracleithrum is ornamented. In the holotype (IGM 6716) this area is garnished with a series of parallel and short ridges, which become gradually more tenuous in dorso-ventral direction ( Figs 6–7 View FIGURE 6 View FIGURE 7 ). In the dorsal portion of the bone, these ridges project as spines beyond the bor- der of the bone. However, the same area is ornamented with tubercles in IGM 6719 ( Fig. 9 View FIGURE 9 ) and with tubercles and ridges in IGM 6720 ( Fig. 8 View FIGURE 8 ).
In lateral view the cleithrum has a crescent shape, slightly deeper than long and anteriorly reaches a level around the posterior rim of the orbit. The cleithrum has up to seven sinuous and variably interrupted series of denticles along the edge between the branchial and lateral surfaces. The lateral surface of the cleithrum is notably narrow and its free border is smooth. The four postcleithra are aligned following the curved made by the posteroventral border of the cleithrum. Differing from the other dermal components of the pectoral girdle, the exposed portion of the postcleithra is covered with a layer of ganoin. The first, most dorsal postcleithrum is dorsoventrally elongate, narrowing dorsally. It is the deepest among the postcleithra, but it is narrower than the adjacent postcleithrum. This second postcleithrum is approximately as deep as long and has a very gently concave anterior border, and a convex posterior border. The two following postcleithra are much smaller than the elements described before. Both are elongate and their shape accommodate to the shape of the posterior border of the cleithrum. As is the case of the supracleithrum, the two most dorsal postcleithra are variably ornamented with tubercles and/or ridges, which are sometimes projected in spines. Ventral and medial to the second postcleithrum there is a modified scale with three strong spines projecting backwards ( Figs 6–7 View FIGURE 6 View FIGURE 7 ). To facilitate comparisons (see discussion) we refer to this scale, which is well exposed in the holotype (IGM 6716), as the modified pectoral scale.
Of the endochondral components of the pectoral girdle, only five radials are exposed in the holotype ( Figs 6– 7 View FIGURE 6 View FIGURE 7 ). Three of these radials are only partially exposed because they are preserved in articulation. The other two radials are disarticulated and displaced ventral to the skull and anterior to the pectoral fin.
Paired fins. The pectoral fin is best and almost completely preserved (only the distal fin rays are missing) in the holotype (IGM 6716; Figs 6–7 View FIGURE 6 View FIGURE 7 ), comprising nine lepidotrichia. The proximal third to one half of each lepidotrichium is unsegmented, contrasting strongly with the distal portion, which is finely segmented and intensively branched. The first lepidotrichium does not branch and its terminal segment is elongate, covered with ganoin and completes the series of fringing fulcra, which are laying on this first ray. The proximal portion of the first pectoral ray forms a lateral ridge, which is also covered with ganoin. Scattered remains of a few small broken spines are preserved on the proximal portion of the second to fourth pectoral rays. From the second ray onwards, the lepidotrichia are asymmetrically branched, the medial portion being always more intensively branched than the lateral one. Also, the branching is gradually more intense towards the medial rays. Two short and paired basal fulcra are preceding the series of short fringing fulcra.
The pelvic fins are also best, though only partially preserved in the holotype (IGM 6716; Fig. 2 View FIGURE 2 ), where the pelvic fin is located about 0.52 of the standard length, right behind the seventh vertical row of scales. The total number of rays is unknown, but at least two lepidotrichia are preserved. Resembling the case of the pectoral fin, the proximal segment of the first pelvic fin ray forms a lateral ridge, which is covered with a thin layer of ganoin. A series of fulcra garnish the pelvic fin, including one unpaired basal fulcrum, one paired basal fulcra, and at least eight fringing fulcra.
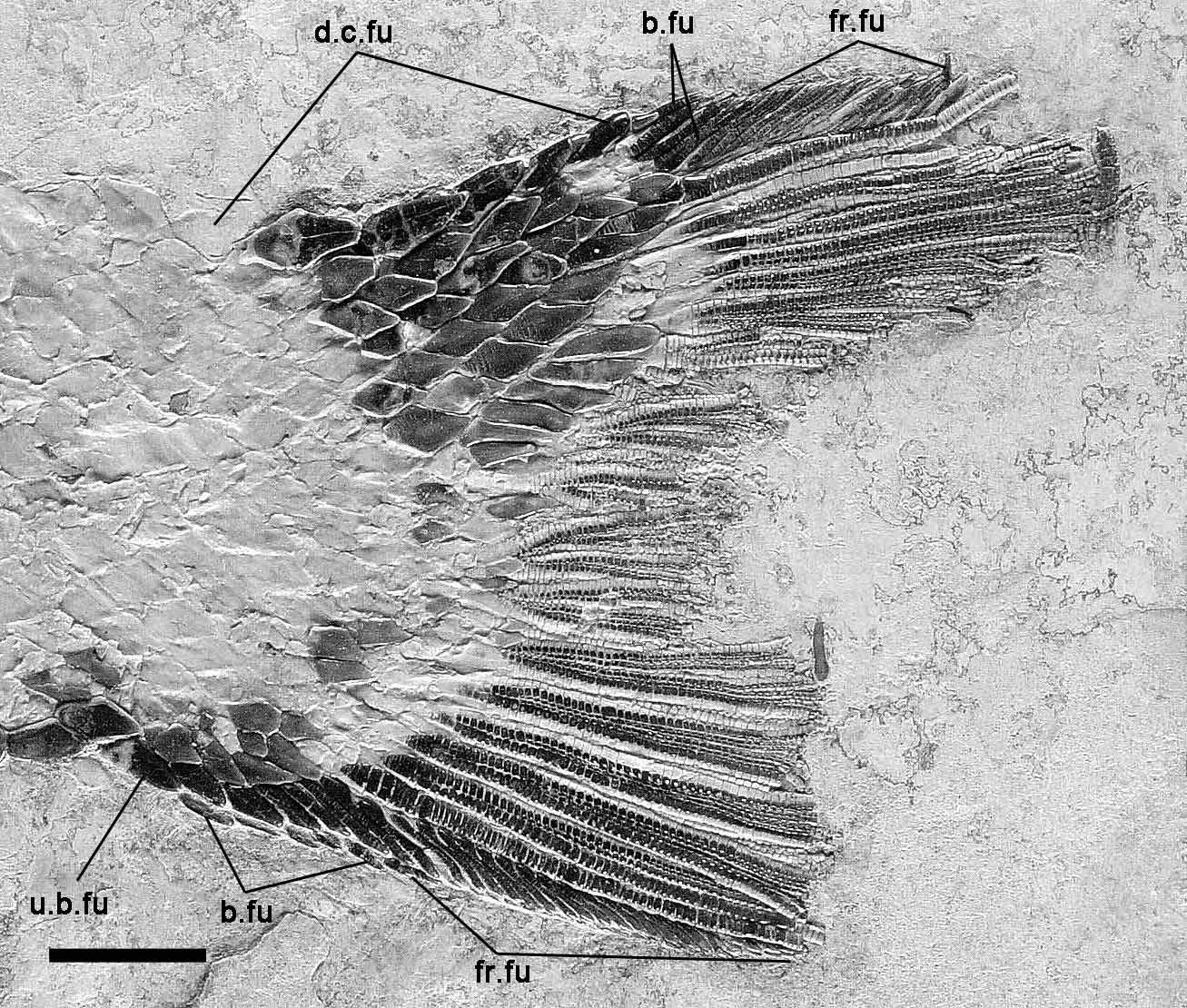
Unpaired fins. The unpaired fins are best preserved in IGM 6718 ( Figs 3 View FIGURE 3 B, 13–14). The dorsal fin is short and originates right back of the level of the pelvic-fin insertion. The dorsal fin is incompletely preserved in IGM 6716 and 6720 ( Figs 2 View FIGURE 2 , 5 View FIGURE 5 ), but both specimens show that this fin originates at the 23rd vertical row of scales, at about 0.6 of the standard length. There are 13 lepidotrichia in the dorsal fin of IGM 6718 ( Fig. 13 View FIGURE 13 A), the height of which decreases progressively posteriorly and, thus, the shape of the fin was probably triangular. All dorsal fin rays, including the first one, branch at least two times. There are five large paired basal fulcra, and at least 10 large fringing fulcra. Among these fulcra, the two posterior paired basal fulcra and two anterior fringing fulcra are the largest, being almost 1.5 times higher than the adjacent fulcra.
The anal fin is also incompletely preserved in both IGM 6716 and 6720 ( Figs 2 View FIGURE 2 , 5 View FIGURE 5 ), but these specimens show its origin at the 18th vertical row of scales, at 0.72–0.74 of the standard length. The anal fin is almost complete and well preserved in IGM 6718 ( Fig. 13 View FIGURE 13 B), comprising seven lepidotrichia, the two distal thirds of which are finely segmented and branched, including the first ray that branches at least once. The proximal half of the first lepidotrichium is covered with lateral patches of ganoin. The anal fin is garnished with three paired basal fulcra, and at least 19 small fringing fulcra. There is a single large preanal scute, which is about three times larger than the adjacent scales.
The caudal fin is also very well preserved in IGM 6718 ( Fig. 14 View FIGURE 14 ), but the distal portions of the fin rays are missing, so that the shape of the fin is unknown. Both in this specimen and in IGM 6720, there are nine and eight lepidotrichia forming the dorsal and ventral caudal lobes respectively. As shown in IGM 6716, all caudal fin rays are laterally covered with patches of ganoin. The caudal fin rays are intensively branched, with the exception of the two marginal rays, and finely segmented like the lepidotrichia in the other unpaired fins. In this case however, the proximal unsegmented portion is short and almost totally covered with scales. As far as preserved, the dorsal and ventral marginal rays are unbranched. The most posterior flank scales are relatively longer and oval. There are at least six unpaired dorsal caudal fulcra covering the body lobe of the tail, followed by three paired basal fulcra and at least 13 fringing fulcra laying on the dorsal marginal ray. Ventrally, the caudal fin is preceded by one short unpaired basal fulcrum and five paired basal fulcra, and there are at least 22 small fringing fulcra on the ventral marginal ray.
Squamation. The body is covered with large and thick rhomboid scales. The squamation pattern is best preserved in the holotype (IGM 6716; Fig. 2 View FIGURE 2 ), in which are 34 vertical rows of scales from the operculum to the hinge line, and a complete ridge of scales along the dorsal midline between the skull and the dorsal fin. The scales are not ornamented and have smooth surfaces. Most of the scales on the flank in IGM 6716 form one or two spines at their posteroventral angle ( Fig. 6 View FIGURE 6 ). However, such spine is only insinuated in the scales of IGM 6719 ( Fig. 4 View FIGURE 4 ) and generally absent in IGM 6720 ( Fig. 5 View FIGURE 5 ), which shows that the feature is highly variable within the species. On the flank, the scales are generally rectangular, approximately as deep as long, only somewhat deeper than long immediately behind the operculum. The dorsal ridge scales are conspicuous, slightly longer than those adjacent scales and bears a short posterior spine directed backward ( Fig. 15 View FIGURE 15 A). In the caudal peduncle, the scales are about 2 times longer than deep and have smooth borders in all the specimens. There are seven inverted rows of scales forming the body lobe of the tail. Among them, the scales of the marginal row are distinctively elongate and not diamond shape as the other scales in the caudal region. Six scales are preserved in this marginal row, but there is a fragment of a seventh scale and one scales is evidently missing both ventrally and dorsally and, thus, nine is the total number of scales in this row. Nevertheless, this marginal row does not reach the tip of the body lobe, which is formed by the two following rows of scales.
The articular processes of the scales are only shown in the middle flank region of IGM 6717. In this area, the scales show well-developed articular, anterodorsal and anteroventral processes (anterior pegs) ( Fig. 15 View FIGURE 15 B). The articular process that serves in the peg-and-socket articulation (dorsal peg) is not very high and places right at the base of the anterodorsal process. The anterodorsal and anteroventral processes serve in the longitudinal articulation and are oriented in diverging directions, with the anterodorsal process being larger than the anteroventral process.










| IGM |
Geological Institute, Mongolian Academy of Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.