
Euphyllia baliensis, Turak, Emre, Devantier, Lyndon & Erdmann, Mark, 2012
|
publication ID |
https://doi.org/ 10.5281/zenodo.215180 |
|
DOI |
https://doi.org/10.5281/zenodo.6180116 |
|
persistent identifier |
https://treatment.plazi.org/id/039A8772-FFCE-AA79-FF08-FE0AE325D1A0 |
|
treatment provided by |
Plazi |
|
scientific name |
Euphyllia baliensis |
| status |
sp. nov. |
Euphyllia baliensis sp. nov.
(Figures 1–3, Tables 1–2)
Material examined. Holotype. MZB CC1520, Padang Bai, Bali, Indonesia, 08° 31’ S, 115° 34’ E, depth 37 m, coll. M.V. Erdmann, 02/05/2011. The holotype consists of a branched piece of a colony of 57 mm total length.
Paratypes. Four at the Indonesian Institute of Sciences' Research Centre for Biology ( MZB), Cibinong, Java, Indonesia: 1— MZB CC1516, 2— MZB CC1517, 5— MZB CC 1518, 10— MZB CC 1519, three at MTQ: 4—G65034, 8—G65035, 9—G65036, one at the Natural History Museum, London, UK: 6– NHMUK 2011.458 and one at the Smithsonian Institution, Washington D.C., USA: 7– USNM 1156955. All paratypes from same location, depth and date as holotype, coll. Emre Turak. Paratypes resemble holotype, ranging in total length from 39 to 79mm.
An additional six specimens are also lodged with MTQ, as G65037, G65038, G65039, G65040, G65041, G65042.
Diagnosis. In the field this species is recognizable as a diminutive type of ‘bubble coral’, growing as low cushions covered by compacted, extended, brown-green coloured fleshy polyps of characteristic ‘anchor – glove’ shaped Euphyllia form. Polyps do not readily retract, but when persistently disturbed will partially retreat in to small calices, revealing thin and delicate branches, usually less than 10cm tall. Branching is mostly in the form of bifurcation. Corallites are small, around 3mm diameter, round and become elongate as they mature to pre-division stage. Septa are not numerous—averaging around 16, in 2–3 cycles and do not extend noticeably beyond the corallite wall rim.
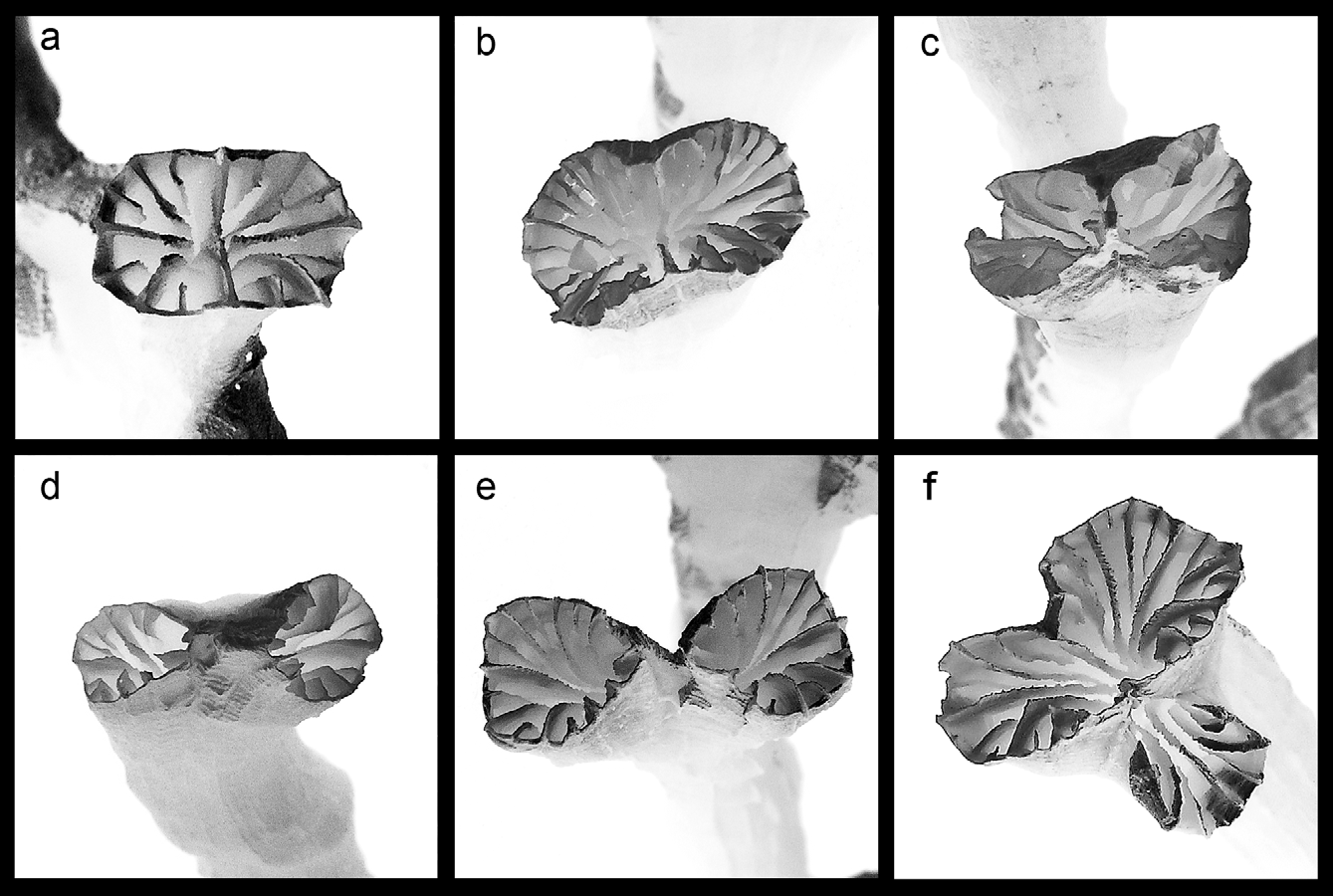
Description. Corallum. Colonies are phaceloid, formed by repeated, dichotomous branching (Fig. 1a, Fig. 2 View FIGURE 2 ), and exhibit indeterminate growth. Mature colonies typically cover an area ranging from ca 200cm 2 to> 1000cm 2, becoming larger monospecific ‘beds’.
Branches are sub-circular and, in the material examined, range from 39–79mm in total length and 2.5– 5mm in diameter, being widest just prior to bifurcation, with new branchlets (up to 4 bifurcations per branch) occurring at 5–25mm intervals along the growth axis (Table 2, Fig. 1a,i).
Branches typically end in a single or pair of corallites, the latter ranging from 2–7mm apart on their proximal sides (Fig. 1a,g,i). These corallite pairs are typically 6–12mm from adjacent corallite pairs, giving a compacted appearance to tops of colonies ( Fig. 3 View FIGURE 3 a,c). Branch extension and colony growth occur via intra-tentacular budding of corallites, initiated by the fusion of one opposing pair of first cycle septa ( Fig. 2 View FIGURE 2 ). This typically results in formation of two daughter corallites, rarely three. The frequency of branching, the distance between branches and branch diameters are relatively uniform within colonies. Colonies are only lightly calcified and very fragile.
Corallites. Corallites are sub-circular, with non-budding corallites averaging 3.1mm diameter and ranging from 2–4.1mm, with very thin walls (Table 2, Fig. 1b–f). Corallites become more elongate prior to budding (Fig. 1e), with minimum and maximum dimensions of 2.8mm and 7.1mm. Septa are widely spaced, of two to three cycles, with the first cycle extending inwards for 2/3 or more of the calice radius (R) before either fusing or plunging vertically or near-vertically to the calice centre. The second cycle typically extends from 1/3–1/2 R and the weakly developed third cycle to 1/5 R (Fig. 1b–f). Inner septal margins are straight to slightly wavy or twisted (Fig. 1f). Septa are smooth to very finely serrated and wavy on their upper margins, with smooth or fine and sparsely granulated sides, and first and second cycles are level with, or extend only slightly (<1 mm) above, corallite rims (Fig. 1a, f–h). The number of septa per corallite ranges from 8 to 30 (avg. 16.65 and se. 1.04).
The septal pattern in mature corallites is based on a tetra or octameral pattern, with four or eight septa in the first and second cycles, and up to 16 septa in an irregular third cycle (Fig. 1b–e). Although unusual, mature corallites in various other Scleractinia also show non-hexameral septal arrangements (J.E.N. Veron, B. Rosen pers. comm.). These include species of Madracis , Cyphastrea , Porites, Stylocoenia, Astrocoenia and Actinacis , the latter three genera all being extinct. The mature corallites of faviid species Cyphastrea microphthalma ( Lamarck, 1816) and poritid Porites decasepta Claereboudt, 2006 have a decameral septal pattern. The hexameral pattern is not considered a basal property of the Scleractinia (J.E.N. Veron pers. comm.), despite the order’s inclusion in sub-class Hexacorallia, which denotes the far more typical occurrence of six-fold symmetry, particularly at initial metamorphosis from the planula larva.
TABLE 2. Measurements of skeletal morphology of holotype and nine paratypes of Euphyllia baliensis sp. nov. Total branch length was measured from base of branch to the tip, length of 2nd branchlet from base of 1st bifurcation to tip, length of 3rd branchlet from base of 2nd bifurcation to tip and so on. For branch diameters, two measurements were taken for the main branch prior to bifurcation, the first at the base of the branch, the second just below the bifurcation. For subsequent branchlets, measurements were taken at the base of the branch above the bifurcation. Corallite diameters were made of mature corallites prior to division and where corallites were not circular, maximum and minimum diameters were taken.
Columellae are absent, a typical feature of the genus (Fig. 1i). Transverse sectioning of branches revealed that first endothecal dissepiments occur at 5–7mm below the corallite rim (Fig. 1i). Endotheca is vesicular, but dissepiments are not numerous and often because of the small diameter of the corallite, traverse the full length of the septal plane to the centre of the calice. They are usually spaced 1mm or less apart.
Costal development is highly variable in height, thickness and length, both within and among colonies. Usually two, occasionally three, orders are present, the second and third orders sometimes occurring as more or less discontinuous lines of granules. Costae usually show maximum development around the mid section of branches. Primary costae may be up to 0.4mm exsert. Costae, particularly those of higher orders, may be located in narrow elongate depressions or grooves, which correspond to septa on the inner side of the wall (Fig. 1a,g,h). Costae are often wavy, particularly on outer corallite walls near branch tips, with undulations becoming less apparent towards the base of branches. There may be some ornamentation on the costal margins and when exert, granules may be present on the sides.
The outer wall is highly ornamented, especially close to branch tips, with extensive granulation showing different patterns of alignment, corresponding to variations in septal development. These patterns may be in the form of herring-bone or concentric ridges between costae (Fig. 1g,h).
FIGURE 1. Corallum shape and corallite, outer wall and endothecal detail of Euphyllia baliensis spec. nov. a —Branch of holotype (MZB CC 1520), 57 mm tall, b —Newly formed corallite from paratype 10 (MZB CC1519). c —Young corallite (2nd stage) from paratype 1 (MZB CC1516). d —Young corallite from holotype. e —Mature corallite from holotype. f —Corallite septal detail from paratype 2 (MZB CC1517). g —Outer wall detail from paratype 5 (MZB CC1518). h —Outer wall detail from paratype 2. i —Cross section from non-type material showing dissepiments and lack of columella. Scale bars; 10mm in 1a and 2mm in 1b-i. Photos by E. Turak.
Polyps. Polyps have numerous (> 18) tubular tentacles extended during the day, protruding for 3–7mm above calices, producing a dense, fleshy cover partly or completely obscuring the polyp mouths and the underlying phaceloid colony form. Tentacles are dull to dark translucent brown with lime green bases. Tentacles are ‘anchor’, ‘kidney’ or ‘hammer’ shaped at their tips, occasionally with additional smaller bulbous protuberances, the latter resembling ‘mittens’ or ‘gloves’ ( Fig. 3 View FIGURE 3 a–c).
Habitat and distribution. This species is currently only known from the type locality of Padang Bai in east Bali, Indonesia. The authors have conducted extensive reef surveys around eastern Indonesia (including 47 other sites in a circumnavigation of the islands of Bali, Nusa Penida and small adjacent islands) but have never recorded this species previously; as such, it may currently be considered to have a very restricted range. It was observed and collected in depths of 27–37 m on a moderate slope exposed to strong currents and frequent cool-water upwellings. The new species occurs there in extensive beds interspersed among the calcareous green alga Halimeda , hydroids, alcyonacean soft corals Dendronephthya sp., and a variety of other deeper water reef corals.
Affinities. The phaceloid corallum morphology is shared with seven other extant members of its genus, although in the present species, the diminutive size of branches and corallites is highly distinctive. Polyp form is most similar to Euphyllia paraancora and to a lesser extent E. ancora , although the occasional occurrence of ‘mitten – glove’ like tentacle tips is also reminiscent of E. divisa and E. yaeyamaensis . Again the relatively very small sizes of polyps and tentacles are distinct.
Remarks. Corallum bases and branches are heavily encrusted by epibenthos, particularly sponges ( Fig. 3 View FIGURE 3 d), such as chalinids and others (Lori Colin, pers. comm.), which may aid in maintaining the structural coherence of colonies in situ. Although depth of the known habitat (> 25m) precludes physical impact by wave energy, nest building by Titan Triggerfish Balistoides viridescens ( Bloch & Schneider, 1801) and other biogenic forms of physical disturbance may promote colony fragmentation; with subsequent localized dispersal assisted by episodically strong current flow.
Etymology. Named for the type locality of Bali, Indonesia.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.