
Epicadus Simon, 1895
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4147.3.4 |
|
publication LSID |
lsid:zoobank.org:pub:BD34D436-7AB1-49F5-AD35-76D91AF3AEEB |
|
DOI |
https://doi.org/10.5281/zenodo.6057215 |
|
persistent identifier |
https://treatment.plazi.org/id/039B87E6-FFC6-9755-7EF3-FC89D63BFADD |
|
treatment provided by |
Plazi |
|
scientific name |
Epicadus Simon, 1895 |
| status |
|
Epicadus Simon, 1895 View in CoL View at ENA
Epicadus Simon, 1895: 1052 View in CoL (Replacement name for Eripus Walckenaer, 1833 View in CoL , preoccupied by Eripus Dejean, 1829 View in CoL [ Coleoptera View in CoL : Carabidae View in CoL ]). Petrunkevitch 1928: 161; Mello-Leitão 1929: 93; Roewer 1954: 755; Brignoli 1983: 604; World Spider Catalog 2016.
Type species. Thomisus heterogaster Guérin-Méneville, 1829 by monotypy.
Composition. Three species: Epicadus heterogaster ( Guérin-Méneville, 1829) ; Epicadus rubripes Mello- Leitão, 1924 and Epicadus planus, Mello-Leitão, 1932 .
Etymology. No etymological explanation was provided in the original description by Simon. Maybe “ Epicadus ” refers to Cornelius Epicadus , servant and biographer of the roman general and statesman Lucius Cornelius Sulla Felix. Epicadus is a masculine anthroponomy from the Southern Illyria (the Roman Dacia). In a similar way, another anthroponomy of the same origin, “ Epidius ” was used by Thorell to name another thomisid genus.
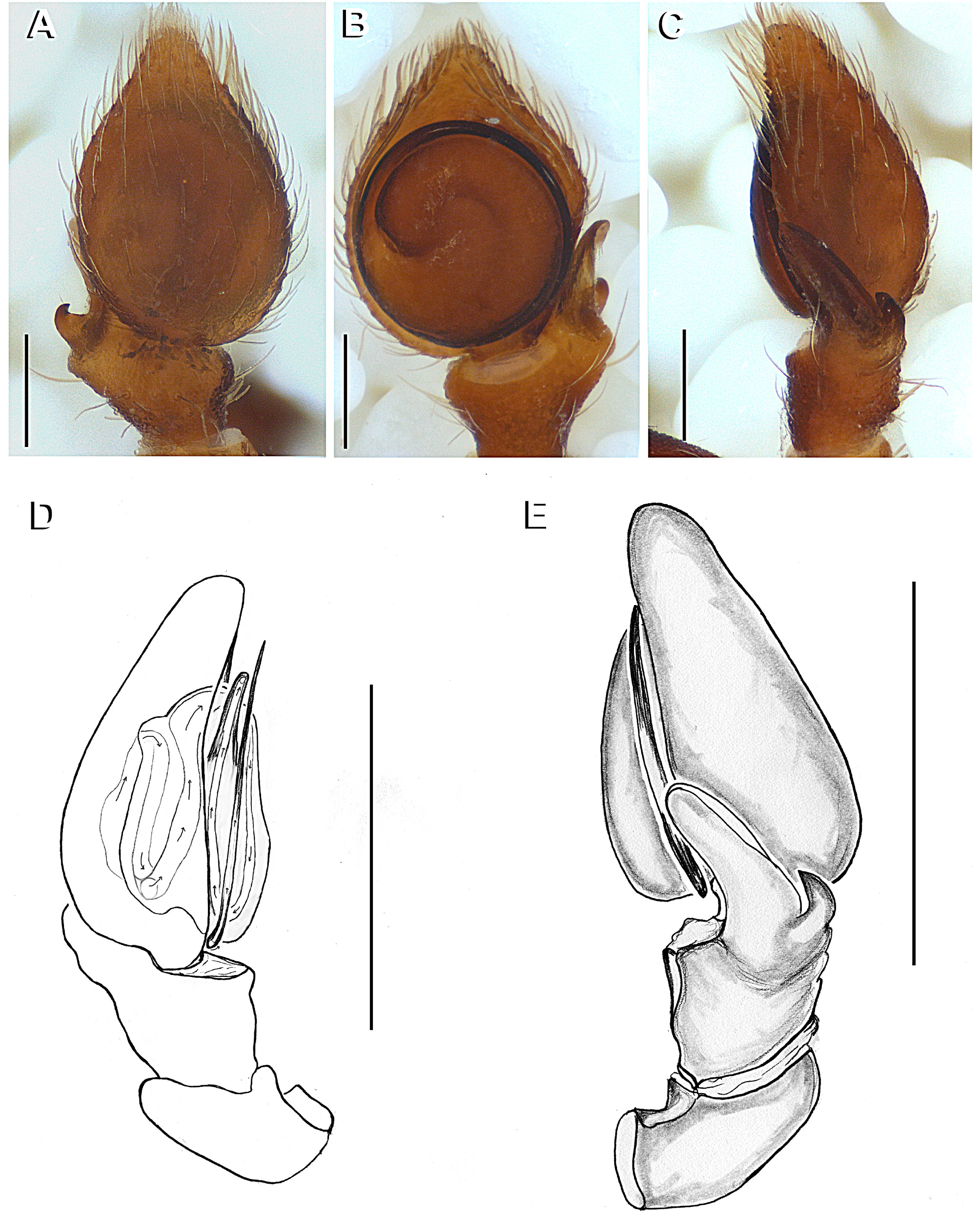
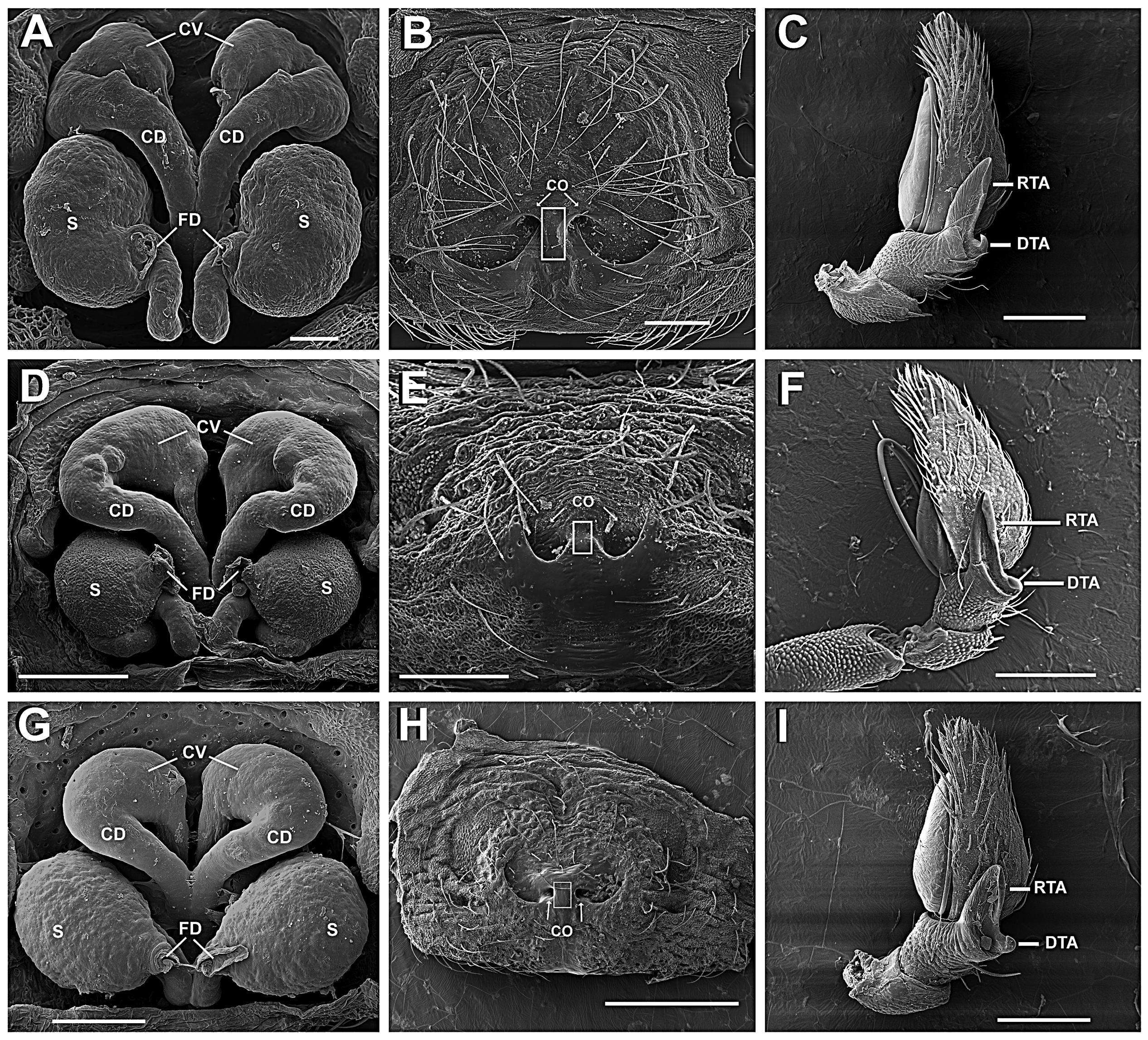
Diagnosis. Epicadus species are distinct from all other Neotropical Thomisidae by having a set of robust and conical opisthosoma projections ( Figs 2 View FIGURE 2 A, 7C, 11C) and a median spire (MS) on the fovea region ( Figs 2 View FIGURE 2 B, 7B, 11B) associated with a white spot. The epigynal plate is glabrous, with a incomplete posterior septum and lower edges, and copulatory orifices sclerotized and located on a C-shaped structure ( Figs 3 View FIGURE 3 A; 8A; 12E). Copulatory ducts S-shaped, with CV about 2–3 times wider that the width of the remaining duct ( Figs 15 View FIGURE 15 A, D, G). Males have a scaly opisthosoma, palp with an elongated and concave RTA sharing its basis with the DTA, whereas DTA is short, acute and curved at the edge ( Figs 5 View FIGURE 5 E, 10D, 14D).
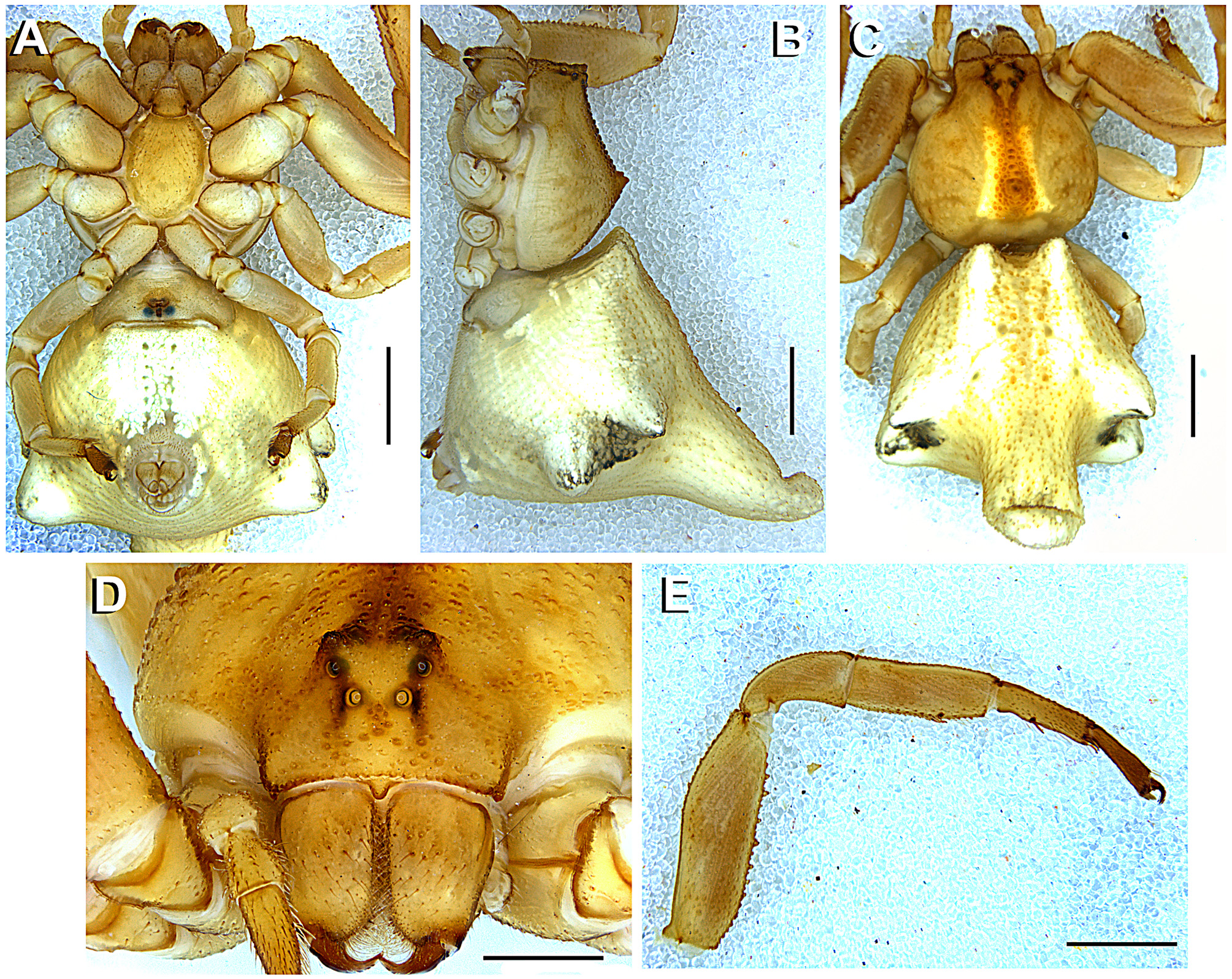
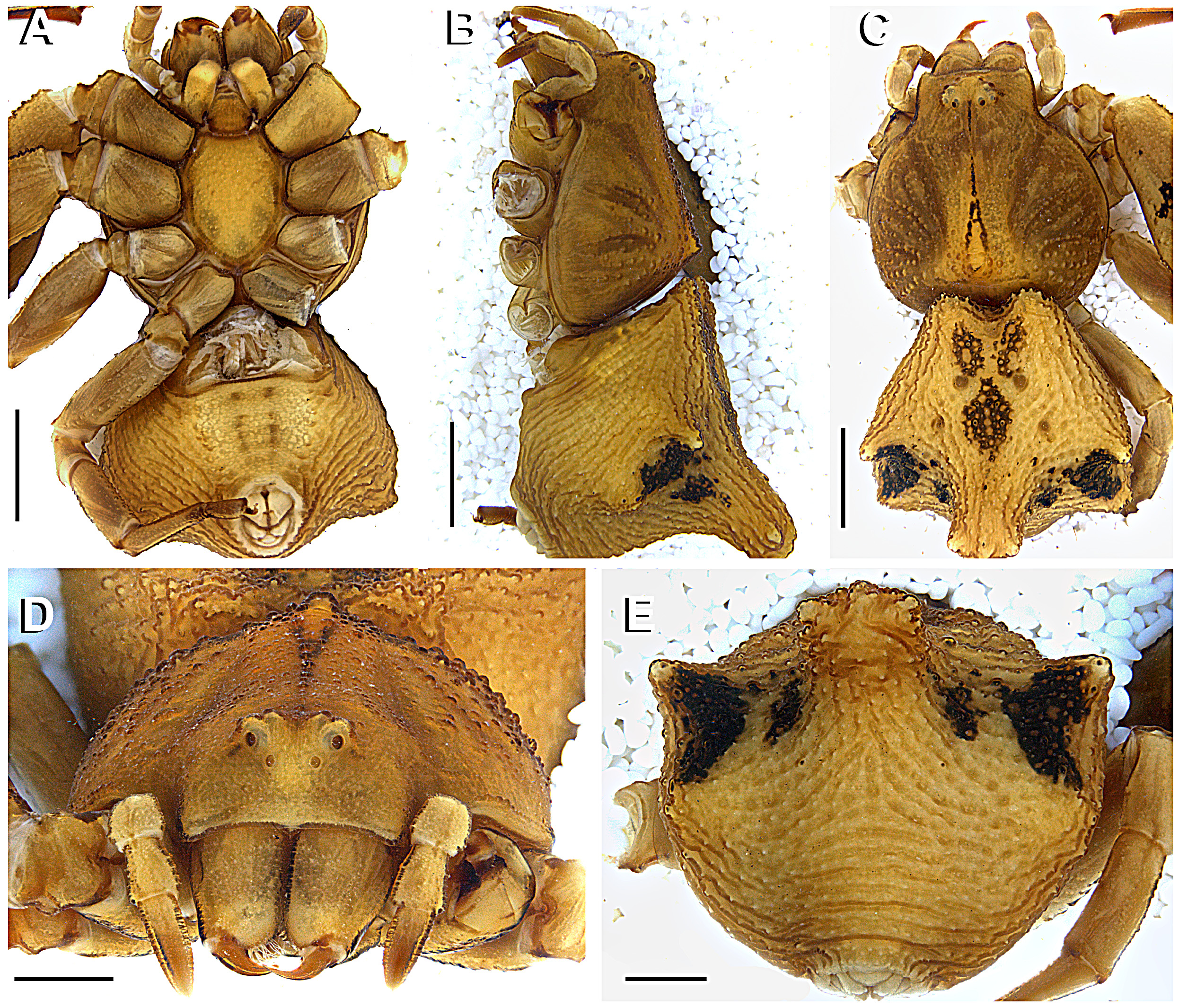
Description. Medium sized spiders, with very marked sexual size dimorphism (total length 2.50–3.70 in males, 10–15 in females). Prosoma as long as wide and higher in the posterior half. Prosoma glabrous, covered with small, colorless granules anteriorly and larger ones in posterior half, along the radial furrows ( Fig. 7 View FIGURE 7 B). Prosoma with a distinct median guanine spot, white in E. heterogaster and E rubripes , or instead a median black stripe or row of black granules ( E. planus ) ( Figs 2 View FIGURE 2 A; 7C; 11C). The female fovea area shows a distinctive median spire, (MS), covered with granules. MS may be discrete ( E. planus , Fig. 11 View FIGURE 11 B) or pronounced ( E. heterogaster , E rubripes , Figs 2 View FIGURE 2 B; 7B). Eyes subequal, arranged in a 4–4 disposition. Eye mound with vertical conical projections, pronounced in E. heterogaster , discrete in E. rubripes and absent in E. planus ( Figs 6 View FIGURE 6 A; 7D; 11D). Chelicerae with three promarginal and two retromarginal teeth; promargin hirsute, with fang basis bearing long and delicate setae. Endites elongated, rounded on the edge, with scarce promarginal scopula. Labium short, truncated, not passing the half of the endites, Sternum oval with concave anterior border. Opisthosoma with distinctive conical abdominal projections distributed as follows: two anterior (absent in males of E. rubripes and E. planus ), two upper lateral, two lower lateral (subtle on males of E. rubripes and E. planus ) and one median posterior. Opisthosoma projections can be directed vertically ( E. heterogaster , Fig. 2 View FIGURE 2 B; median posterior projection in E. rubripes , Fig. 7 View FIGURE 7 B) or horizontally (anterior and lateral projections in E. rubripes , Fig. 7 View FIGURE 7 B; and E. planus — Fig 11 View FIGURE 11 B). The anterior projections might be distinctive, conical ( E. heterogaster , Fig. 4 View FIGURE 4 C) or subtle, like an invagination of the anterior margin of the opisthosoma, slightly flattened dorsoventrally ( E. rubripes ; E. planus , Figs 7 View FIGURE 7 C, 11C). Male opisthosoma covered with small sclerites, darker than the overall opisthosoma color. Spinnerets surrounded by a sclerotized ring ( Fig. 13 View FIGURE 13 E).
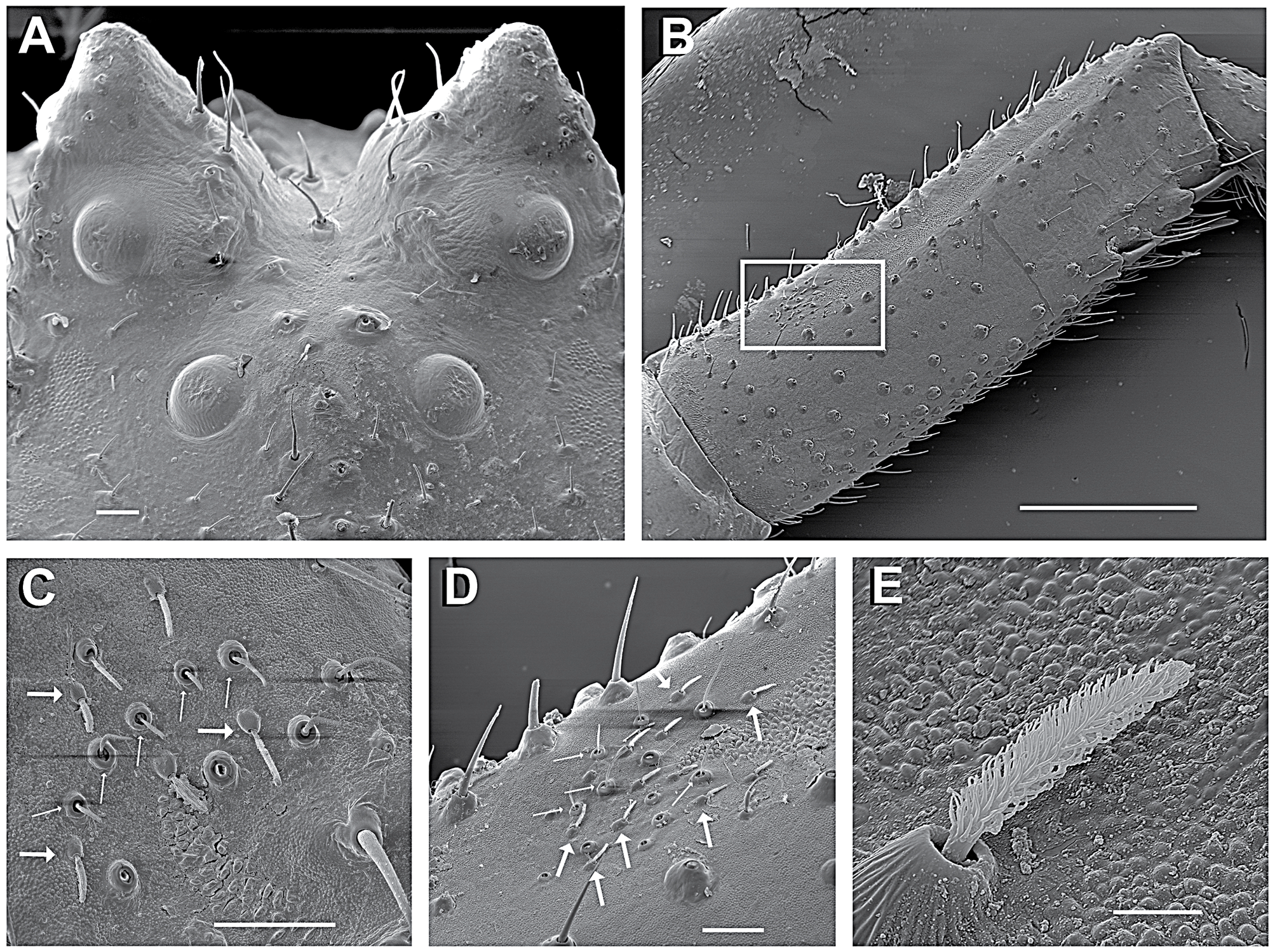
Anterior legs laterigrade and superposed (leg I laying over leg II when spider in resting position, Fig. 1 View FIGURE 1 ); leg III anteriorly oriented and leg IV posteriorly oriented. Anterior legs distinctively larger and stouter than the posterior ones. Femora I and II armed with two rows of ventral conical tubercles (FeT, Figs 2 View FIGURE 2 E, 7E), patellae I and II elongated. Tibiae and metatarsi I and II round in cross-section, armed with short, slightly curved macrosetae, with rounded base and acute edge (sometimes bifid, Fig. 3 View FIGURE 3 E). Metatarsi III and IV round in cross-section with distinct setae tuff on the distal third. Anterior tibiae with a concave area on the dorso-retrolateral face ( Fig. 6 View FIGURE 6 B), where a sensorial pit can be found (SPt, Figs 6 View FIGURE 6 C, D) in the proximal part. The same region contains both duster-shaped setae ( Fig. 6 View FIGURE 6 E) and trichobothria ( Figs 6 View FIGURE 6 C, D). Tarsi I and II of similar size as metatarsi, wider near joint to metatarsi. Tarsi III and IV small and rounded in cross-section, forming an acute angle with the metatarsi, with a dense field of setae along its extension. Leg formula I-II-IV-III. Palp with spatulated setae and a tarsal claw. Epigynum with a discrete, colorless incomplete median septum and lowered median area (better seen on SEM, Figs 15 View FIGURE 15 B, E, H). Epigynal plate at the same level as the opisthosoma ( E. heterogaster , E. rubripes ) or slightly elevated ( E. planus ). Copulatory openings sclerotized ( Figs 3 View FIGURE 3 A, 8A, 12A); vestibular edges sclerotized, forming a smooth and rounded atrium ( Figs 15 View FIGURE 15 B, E, H). Copulatory ducts long, S-shaped ( Fig. 3 View FIGURE 3 C) with upper (anterior) curve (CV) enlarged, similar to a secondary spermatheca (see Bonaldo 2000; Machado et al. 2015) at the distal part ( Figs 15 View FIGURE 15 A, D, G) and getting narrower near the copulatory openings and distant from the spermatheca. Spermathecae rounded or bean-shaped. Fertilization ducts long and thin, oriented dorsally near the spermatheca and turning retrolaterally ( Figs 3 View FIGURE 3 C–D, 8B–C, 12B).
Male palp with tibia robust, wider than long, usually bearing colorless setae. Cymbium pyriform and hirsute. Tegulum discoid, without apophyses ( Figs 5 View FIGURE 5 B, 10E, 14E); RTA canoe-shaped, with the concavity facing retrolaterally ( Figs 5 View FIGURE 5 E, 10D, 14D); smaller edge curved to the inner side of concavity. DTA short, acute, curved and sharing the same basis with RTA ( Figs 15 View FIGURE 15 C, F, I). Embolus filiform ( Figs 5 View FIGURE 5 D, 10E, 14E).
Coloration in ethanol (but see comments under “Natural history”): females with yellow or dark yellow prosoma and pearl white opisthosoma. Legs I and II usually bearing black stripes or spots on the dorsum of the femora I and II or (rarely) on metatarsus. Legs and tarsal palp usually black ( Figs 2 View FIGURE 2 , 7 View FIGURE 7 , 11 View FIGURE 11 ). Males are reddish or darkish brown, some specimens dark orange, with very little variation ( Figs 4 View FIGURE 4 , 9 View FIGURE 9 , 13 View FIGURE 13 ). Male opisthosomal sclerites vary between dark orange to dark brown.
Distribution. The genus Epicadus has a classic Neotropical distribution pattern (sensu Morrone 2001) ranging from southern Brazil to central Mexico ( Fig. 17).
Natural history. Knowledge about the ecology and behavior of Epicadus species is very scarce. As many thomisids, Epicadus are usually found on leaves or flowers of different plants, where they hunt by the sit-and-wait behavior, and use their strong anterior legs to catch and hold prey ( Figs 16 View FIGURE 16 C, E).







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Epicadus Simon, 1895
| Silva-Moreira, Thiago Da & Machado, Miguel 2016 |
Epicadus
| Brignoli 1983: 604 |
| Roewer 1954: 755 |
| Mello-Leitao 1929: 93 |
| Petrunkevitch 1928: 161 |
| Simon 1895: 1052 |