
Anthothela vickersi ( Benham, 1928 ) Moore & Alderslade & Miller, 2017
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4304.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3D557C94-0783-4C39-80C3-9C321DA94800 |
|
DOI |
https://doi.org/10.5281/zenodo.6015365 |
|
persistent identifier |
https://treatment.plazi.org/id/E21199B8-6E49-45C3-BD6E-B85BA74FA46E |
|
taxon LSID |
lsid:zoobank.org:act:E21199B8-6E49-45C3-BD6E-B85BA74FA46E |
|
treatment provided by |
Plazi |
|
scientific name |
Anthothela vickersi ( Benham, 1928 ) |
| status |
comb. nov. |
Anthothela vickersi ( Benham, 1928) View in CoL new combination
( Figs. 20–31 View FIGURE 20 View FIGURE 21 View FIGURE 22 View FIGURE 23 View FIGURE 24 View FIGURE 25 View FIGURE 26 View FIGURE 27 View FIGURE 28 View FIGURE 29 View FIGURE 30 View FIGURE 31 )
Spongioderma (?) vickersi Benham, 1928: 81 View in CoL –83, Figs. 25–31 View FIGURE 25 View FIGURE 26 View FIGURE 27 View FIGURE 28 View FIGURE 29 View FIGURE 30 View FIGURE 31 .
Homophyton vickersi ( Benham, 1928) View in CoL : van Ofwegen et al. 2000–2007 http://www.marinespecies.org/ aphia.php?p=taxdetails&id=288708 accessed May 2017.
Material examined. Holotype: OMNZ IV8728, 21 miles north of Doubtless Bay , east coast of the North Island, New Zealand, H.M.C.S. Iris , col. Lieut. Vickers, depth 945 m, 1926–1928.
Other material: TMAG K3984, S-Tasmania Slope (~ 1000 m), Huon Commonwealth Marine Reserve (CMR), SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 9, sample 23 (SS 200702 /009-023), 44.154– 44.162°S, 147.128– 147.131°E, depth 800–920 m, 31st March 2007 GoogleMaps ; TMAG K3985, Hill U seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 15, sample 38 (SS 200702 /015-038), 44.322°S, 147.181– 147.185°E, depth 1100–1200 m, 1st April 2007 GoogleMaps ; TMAG K3986, Z9 Seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 58, sample 53 (SS 200702 /058-053), 44.202– 44.199°S, 147.318– 147.320°E, depth 1020–1100 m, 7th April 2007 GoogleMaps ; TMAG K4264, Dory Hill seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 49 (SS199701/49), 44.322– 44.34°S, 147.115– 147.072°E, depth 1167 m, 29th January 1997 GoogleMaps ; TMAG K4109, Hill J 1 seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 40 (SS199701/40), 44.243– 44.273°S, 147.36– 147.323°E, depth 1024–1548 m, 27th January 1997 GoogleMaps ; TMAG K4110, U (R) Hill seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 41 (SS199701/41), 44.318°S, 147.115°E, depth 1314 m, 28th January 1997 GoogleMaps ; TMAG K4111, Dory Hill seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 48 (SS199701/48), 44.313– 44.34°S, 147.142– 147.073°E, depth 1456 m, 29th January 1997 GoogleMaps ; TMAG K4112, Hill A 1 Reserve seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 62 (SS199701/ 62), 44.328– 44.322°S, 147.268– 147.325°E, depth 1261–2253 m, 30th January 1997 GoogleMaps ; TMAG K4113, Hill V seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 67 (SS199701/67), 44.393– 44.387°S, 147.147– 147.23°E, depth 1614 m, 31st January 1997 GoogleMaps ; TMAG K4114, Hill V seamount, Huon CMR, SW Tasman Sea, Australia, CSIRO RV Southern Surveyor, stn. 69 (SS199701/69), 44.397– 44.398°S, 147.147– 147.178°E, depth 1262–1854 m, 31st January 1997 GoogleMaps ; TMAG K4265, Mongrel seamount, Huon CMR, SW Tasman Sea, Australia, stn. J2-386-06, sample 0 1, 44.254°S, 147.115°E, depth 982 m, ROV Jason deployed from the U.S. RV Thomas T. Thompson, team led by Dr Jess Adkins & Dr Ron Thresher, 23rd December 2008 GoogleMaps ; MNHN IK-2009- 535, Ile Amsterdam, Southern Indian Ocean, N/O Marion Dufresne, stn. 2 (JASUS, MD50, stn.2, CP07), 37.783°S, 77.65°E, depth 940–1680 m, 9th July 1986; MNHN IK-2009-534, Ile Saint Paul, Southern Indian Ocean, N/O Marion Dufresne, stn. 23 (JASUS, MD50, stn.23, CP113), 38.917°S, 77.633°E, depth 1065–1125 m, 19th July 1986; NIWA 40439 View Materials , Macquarie Ridge, NE of Macquarie Island, Southern Ocean , NIWA RV Tangaroa , stn. 77 ( TAN 0803 View Materials /77), 53.738°S, 159.114°E, depth 1014– 925 m, 11th April 2008 GoogleMaps ; NIWA 40508 View Materials , Macquarie Ridge, NE of Macquarie Island, Southern Ocean , NIWA RV Tangaroa , stn. 79 ( TAN 0803 View Materials /79), 53.715°S, 159.131°E, depth 770– 810 m, 12th April 2008 GoogleMaps ; NIWA 40578 View Materials , Macquarie Ridge, NE of Macquarie Island, Southern Ocean , NIWA RV Tangaroa , stn. 81 ( TAN 0803 View Materials /81), 53.731°S, 159.166°E, depth 1150–1270 m, 12th April 2008 GoogleMaps ; NIWA 41129 View Materials & NIWA 41865 View Materials , Macquarie Ridge, S of Macquarie Island, Southern Ocean , NIWA RV Tangaroa , stn. 118 ( TAN 0803 View Materials /118), 59.048°S, 158.901°E, depth 1400–1615 m, 19th April 2008 GoogleMaps ; NTM CO12800, North Macquarie Ridge, NE of Macquarie Island, Southern Ocean , CSIRO RV Southern Surveyor, stn. 97 (SS199901/97), 53.932°S, 159.098°E, depth 364 m, 26th January 1999 GoogleMaps ; NTM CO12801 (ex TMAG K1398), North Macquarie Ridge, NE of Macquarie Island, Southern Ocean , CSIRO RV Southern Surveyor, stn. 120 (SS199901/120), 53.645°S, 159.158°E, depth 1050 m, 30th January 1999 GoogleMaps .
Description:
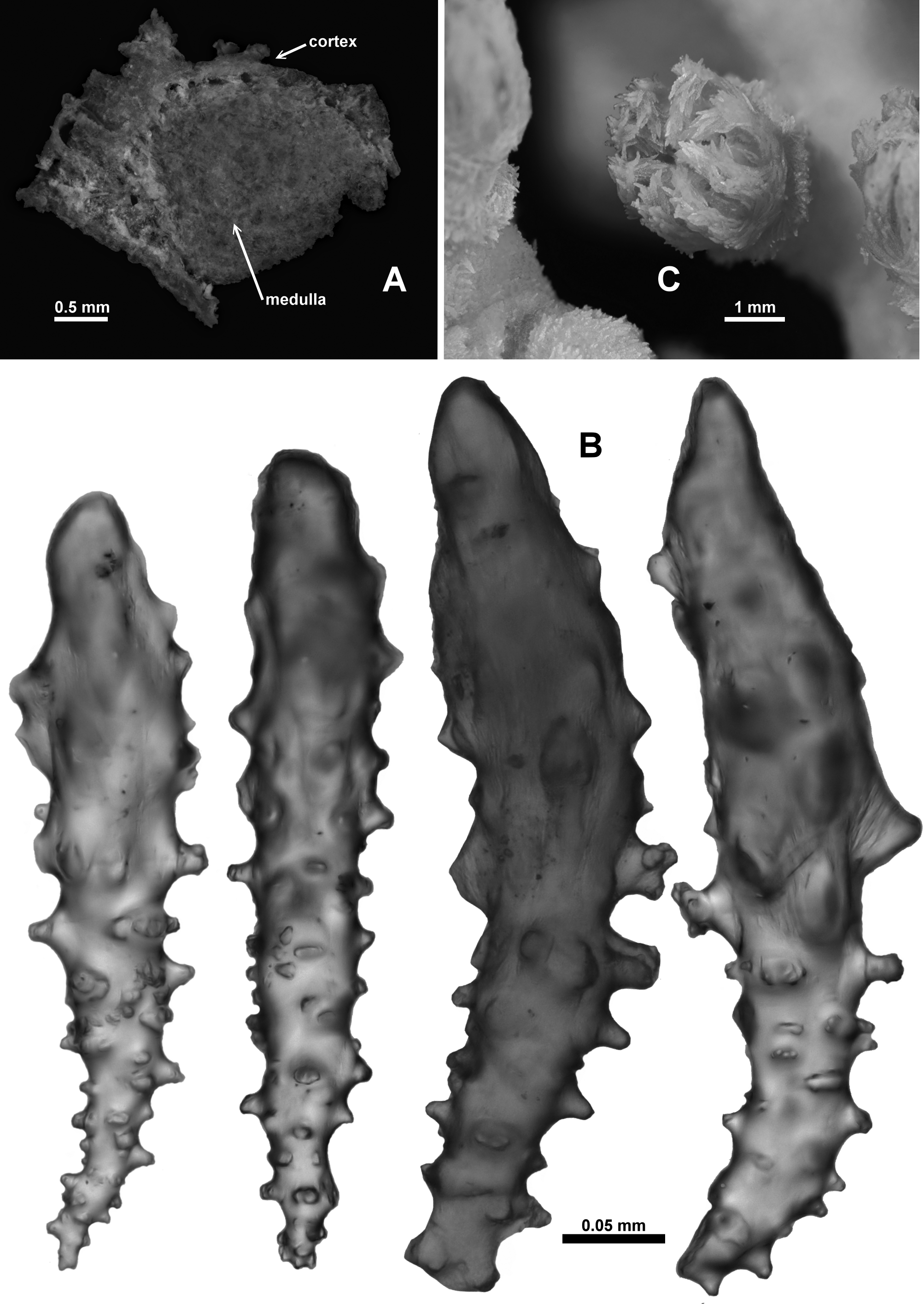
Colony form: The holotype is a piece of a larger colony, and is approximately 60 mm long and 30–40 mm in diameter ( Fig. 20 View FIGURE 20 A, B). There is a sharp bend approximately in the middle of the branch. Benham (1928) states that the colony “had been put into a phial too narrow and too small for it, so that the stem is bent and some of the branchlets are broken off”. There are one or two small circles where the cortex of the colony is missing and the medulla is visible—these may be the origins of the branchlets mentioned by Benham, although they could also be evidence of broken polyps ( Fig. 20 View FIGURE 20 Ba). One end of the main branch is slightly flattened and broadened where there once was a side branch ( Fig. 20 View FIGURE 20 Bb). There are two small side branches remaining, each emanating close to the sharp bend in the colony. One is 13.3 mm long and 3.3 mm wide ( Fig. 20 View FIGURE 20 Aa) while the other is simply a polyp bunch and is 10 mm long (including the length of the terminal polyp) and 11.5 mm wide (which includes two transversely arranged polyps) ( Fig. 20 View FIGURE 20 Ab). The main branch is approximately circular in cross-section although slightly distorted at bifurcation points where it tends to be elliptical. There is no evidence of membranous parts of the colony or anastomoses.
One side of the colony appears to be clear of polyps but this is an artefact of the storage conditions where the colony was secured to a glass slide for many years. Most of the polyps which were on that side of the colony have been bent aside or broken off ( Fig. 20 View FIGURE 20 A cf. Fig. 20 View FIGURE 20 B). The calyces are arranged on all sides without order. They occur close together throughout the colony, with the largest distance between polyps at 4 mm, and are particularly crowded or clumped at the tip of the side branches with no free surface between them.
Grossly, the colony is in reasonably good condition with approximately 30 intact polyps and 18 empty calyces. The surface of the colony is mostly complete and undamaged.
Colour: According to Benham, the colony was “a pale colour” but he is clearly describing it sometime after preservation given he mentions the distortion of the colony due to limited space in the “phial”. It is now light beige in alcohol.
Polyps and Calyces: Calyces are mostly cylindrical, sometimes taller than wide, and are usually at right angles to the branches except for those at the tip which tend to extend at oblique angles from the branch ( Fig. 21 View FIGURE 21 A) and they range from 2.3–4.5 mm high and 2.4–3.5 mm wide. The exsert part of the polyps varies from approximately 2.2–4.5 mm tall; most are partly retracted so the polyp head rests on the lip of the calyx ( Fig. 21 View FIGURE 21 B) but one or two polyps are somewhat more extended so the polyp neck is partly visible ( Fig. 21 View FIGURE 21 C). There are no fully retracted polyps although there is one polyp which only projects very slightly. Tightly crowded sclerites occur throughout, with those on the polyp head arranged as points and a collaret and those on the calyces having the tips projecting out from the surface, giving the calyces a prickly appearance ( Fig. 21 View FIGURE 21 B, C). The tentacles in most of the polyps are folded over into the polyp mouth and so form a rounded mound with eight furrows. According to Benham there are 4–6 “long and narrow” pinnules along each side of the tentacles.
Medulla and Cortex: The branches are composed of a thin cortex surrounding a medulla composed of sclerites tightly packed together. Sclerites are arranged longitudinally to obliquely in both the medulla and cortex. In a cross-section of the axis, a ring of longitudinal boundary canals distinctly separates the cortex from the medulla ( Fig. 22 View FIGURE 22 A). These canals run parallel and close to one another but do not appear to frequently anastomose. Therefore they do not form a true boundary space. There are no obvious coelenteric canals occurring in the central medulla. Two small cross-sections were made from one cut end of the central branch only, so there was no opportunity to assess the occurrence of central coelenteric canals throughout the colony. In one of the crosssections, in the centre of the medulla the density of the sclerites thinned slightly giving an indistinct impression of a central canal. Benham (1928) describes and depicts the boundary canals and later states “it [the medulla] presents no canals, but a few small sub-circular spaces occur towards the centre, which I take to be due to spicules having dropped away”.
Sclerites: The majority of the sclerites are in good condition; however those that project out from the colony (mostly from the calyx and the top of the points) have a smooth and rounded tip. On close investigation, these predominately clavate sclerites have a slight fuzziness around the perimeter of their projecting tip and the tip is brown while the remainder of the sclerite is clear ( Fig. 22 View FIGURE 22 B). Sclerites which do not project from the surface do not have this feature. Given that other species have thorny sclerites of similar size and shape projecting from the calyx and points, it seems most probable that the projecting tip of these sclerites was originally thorny but the ornamentation has been eroded, leaving only slight rounded isolated bumps, perhaps due to a short immersion postmortem in an acidic medium. The damaged sclerites are figured, however caution is necessary when comparing with sclerites from other Anthothela species.
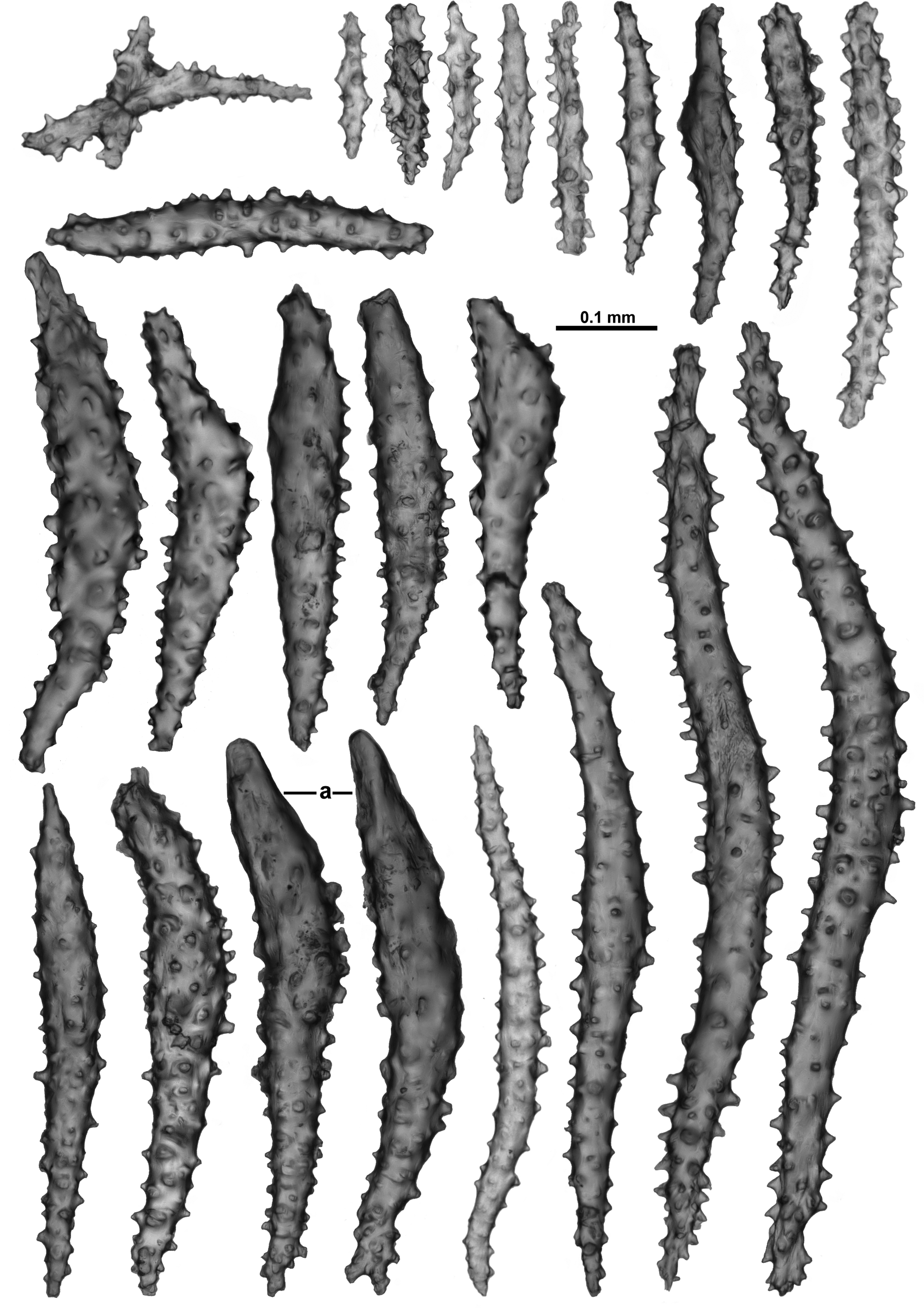
The collaret consists of many transversely arranged sclerites which grade en chevron up the points to be longitudinal in the distal part with many sclerites bunched together and projecting slightly past the base of the infolded tentacles ( Figs. 21 View FIGURE 21 B; 22C). Point and collaret sclerites consist chiefly of long, narrow, straight or slightly curved sticks and spindles with relatively simple tubercles, sometimes sharp ( Fig. 23 View FIGURE 23 ), and warty thorn clubs, some with a smoothed tip ( Fig. 23 View FIGURE 23 a). The simple spindles range in length from 0.11–0.96 mm while the thorn clubs are 0.27–0.56 mm long. There are also rare small crosses and spindles with tubercles in girdles arranged amongst the larger sclerites.
Between the well-defined points, a small number of intermediate sclerites occur, arranged in a narrow line with the ends of the sclerites slightly overlapping each other ( Fig. 21 View FIGURE 21 Ba). They are narrow, tuberculate spindles similar to those found in the points.
The aboral face and the sides of the tentacles are covered in a dense layer of crowded sclerites all arranged longitudinally or obliquely ( Fig. 22 View FIGURE 22 C) with some sclerites projecting above the surface, giving the tentacles a prickly appearance. Most of the tentacle sclerites are short, straight rodlets with simple, relatively sparse tubercles ( Fig. 24 View FIGURE 24 A). Some of the sclerites have a slight curve at one tip where they curve around the side of the tentacles. Most of the rachis sclerites range in size between 0.1 and 0.4 mm long but there are also very small forms, with tubercles mainly in girdles, that appear to be from the distal end of the tentacles where they tend to jumble together and mix with the sclerites in the pinnules. Additionally there are rare, short, thorny clubs ( Fig. 24 View FIGURE 24 B) which are probably from the base of the tentacle rachis and resemble the upper points—these are approximately 0.15–0.46 mm long.
The pinnules are packed with many long, narrow spatulate clubs and sticks and spindles arranged longitudinally with the spatulate end of the clubs in the distal tip of the pinnules ( Fig. 25 View FIGURE 25 ). The spatulate tips are sometimes not well-developed, with some sclerites lacking a markedly splayed end ( Fig. 25 View FIGURE 25 a). Sizes ranges from 0.2–0.52 mm long with no obvious order to the arrangement of the different sizes within the pinnules. There are also small sticks and spindles in the pinnules ( Fig. 25 View FIGURE 25 b) which range from approximately 0.08–0.25 mm long. There are long, narrow sticks and spindles, which are usually straight, with sparse tubercles ( Fig. 25 View FIGURE 25 c), and range from 0.3–0.6 mm long. These appear to extend down into the pinnules but can also be concentrated along the sides of the tentacles.
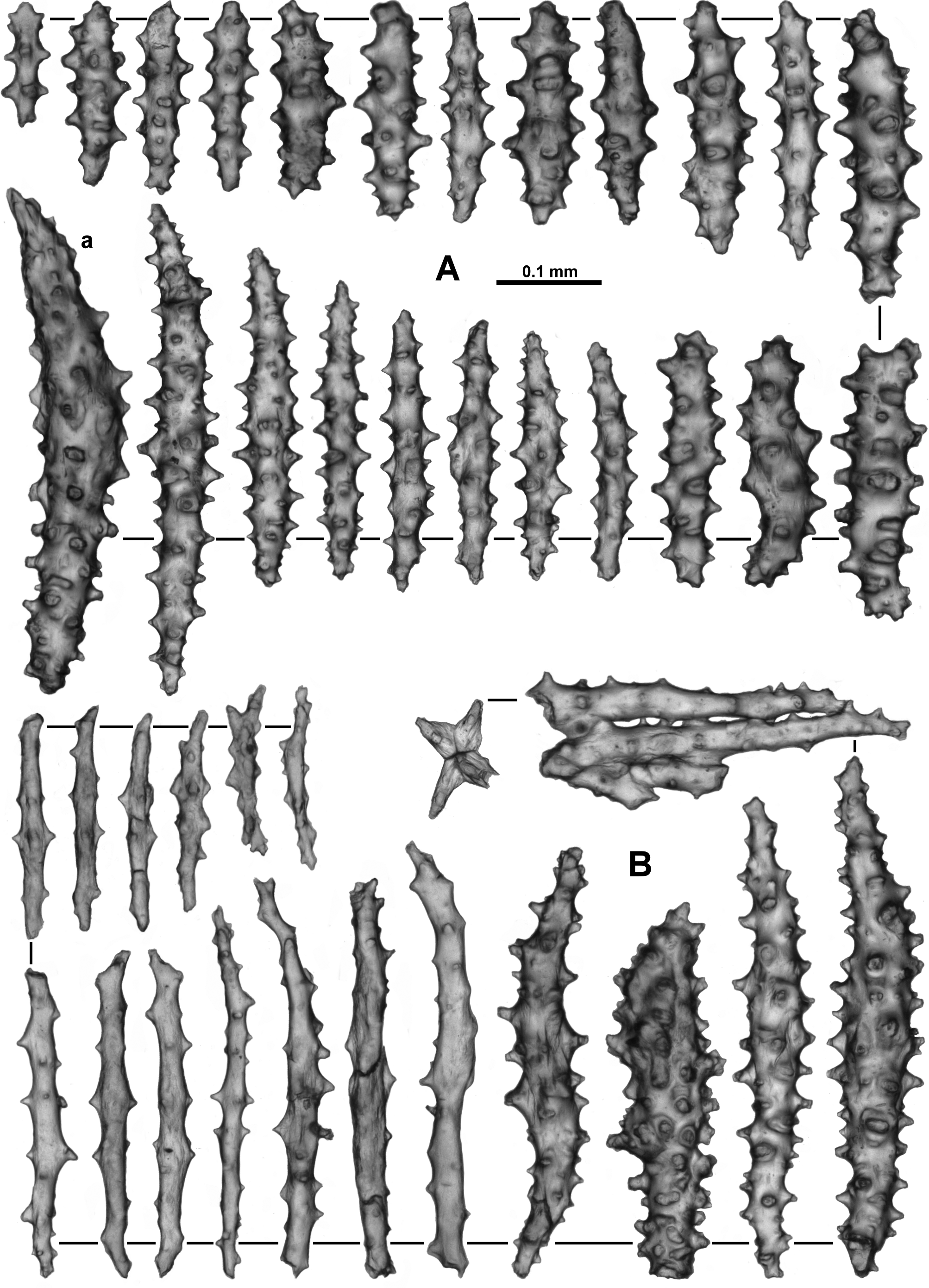
In the pharynx there are small spindles and rods with simple tubercles usually arranged in girdles ( Fig. 26 View FIGURE 26 A). Most of these sclerites are 0.04–0.20 mm in length. They are not crowded and tend to be grouped in indistinct longitudinal lines in the pharynx which correspond to the insertion of the mesenteries ( Fig. 26 View FIGURE 26 B).
The calyces are covered in a dense layer of sclerites, many of which project from the surface giving the calyces a prickly appearance ( Fig. 21 View FIGURE 21 B). A large proportion of the sclerites from the calyx are thorn clubs with an exposed, irregular tip which often appears to be damaged or partially dissolved as mentioned previously ( Fig. 27 View FIGURE 27 a). The latter forms appear to have unaffected warts and tubercles on the embedded portion with some of these being quite complex. Most of the clubs are slightly curved and are 0.29–0.62 mm long. There are other sclerites also present in the calyx, seemingly unaffected, which include many sticks and spindles, 0.18–0.6 mm long, with numerous tubercles, some smaller, clubbed sclerites (0.28–0.33 mm long) and short spindles with tubercles in indistinct girdles (0.11–0.18 mm long; Fig. 27 View FIGURE 27 b).
The cortex has rough, knobbly patches created by densely occurring tuberculate sclerites ( Fig. 21 View FIGURE 21 A). The sclerites are mostly short, stout tuberculate rods and longer, narrower spindles ( Fig. 28 View FIGURE 28 A). The short rods are approximately 0.12–0.28 mm long while the spindles are from 0.2–0.5 mm long. In the only cortex sample taken, a single sclerite was found which could be considered a true thorn club, apparently undamaged ( Fig. 28 View FIGURE 28 Aa). Benham (1928) states the surface “presents a coating of obliquely and irregularly-disposed, short, knobby spicules, very densely fitted together, and below them occur longitudinally-disposed, long, thorny spindles.” This layering may simply be the cortex and medulla.
Sclerites from the medulla also tend to be of two forms; sparsely tuberculate sticks and warty spindles ( Fig. 28 View FIGURE 28 B). The small sample of medulla taken for this re-description has many broken sclerites so their length is difficult to estimate, however Benham (1928) described “long spindles with thorny outgrowths and, the short knobbly rods”. Here the (mostly unbroken) warty spindles range from 0.2–0.5 mm long while the (mostly broken) sparsely tuberculate sticks are 0.13–0.44 mm long. It is likely the tuberculate sticks can be much longer. There is evidence of fusing and branching of the sclerites.
The sclerites are all translucent and colourless under transmitted light except for the damaged tips of the projecting sclerites which are brown.
Variability: The apparent degradation of the tip of the thorn clubs in the points and calyx of the holotype hinders a definitive diagnosis of the species but given factors such as the gross morphology of the colony, general sclerite architecture and geographic proximity, many specimens are here determined as conspecific with the holotype. The assumption is that the rounded tips are an anomaly restricted to the holotype, and the clubs would have once resembled the thorn clubs found in recently collected specimens presented here. If future fresh samples from the type location have similar rounding of the thorn clubs it may be considered a natural feature of the species and determinations herein should be reassessed.
Most of the specimens examined are reasonably consistent with the holotype. In all the samples, long, narrow sticks are arranged along the sides of the tentacles and extend into the pinnules where long spatulate clubs are longitudinally arranged ( Fig. 29 View FIGURE 29 Aa). These sticks are often more frequent than in the holotype but otherwise the tentacle sclerites have a similar form ( Fig. 29 View FIGURE 29 Ba, C). All specimens have thorn clubs in the points ( Fig. 30 View FIGURE 30 A) and calyx ( Fig. 30 View FIGURE 30 B) with more developed thorns and spines on their pointed tip compared with the damaged tip of those in the holotype. Additionally, in the Tasmanian seamount samples, the short, tuberculate rods in the cortex often have one clubbed or pointed, smooth tip ( Fig. 31 View FIGURE 31 Aa) which is not common in the small sample taken of the holotype surface.
Some of the specimens have a large number of long, well-developed thorn clubs in the cortex. The small sample of the holotype cortex had only a single large thorn club but a larger sample may have revealed more given the prickly appearance of the colony surface.
Photographs taken shortly after collection of specimens from the Tasmanian seamounts suggest the live colour is light creamy pink ( Fig. 31 View FIGURE 31 B, C). The Tasmanian seamount specimens are all relatively small suggesting the species does not grow particularly large. Photographs taken on the Tasmanian seamounts suggest it is a reasonably common species, with many colonies of what is presumed to be A. vickersi n. comb. (based on quantity collected, light creamy pink colour and general colony form).
Distribution: Northern New Zealand and the seamounts south of Tasmania, Australia; Southern Indian Ocean.
Depth: 800–1600 metres.
Remarks: When discussing the holotype, Benham acknowledged the lack of relevant literature available to him and his sparse knowledge of the group, and admitted he placed the specimen in the genus Spongioderma “with much hesitation”. He in fact included a question mark in the original designation (“ Spongioderma (?) vickersi ”). It has remained in this now defunct genus mainly due to lack of any comprehensive review of the related taxa and an uncertainty as to the whereabouts of the holotype. Fortunately, the holotype could be located and re-described and it can now be confirmed as a species of the genus Anthothela , based on the spatulate clubs crowded in the pinnules, boundary canals separating the cortex from the medulla, no or indistinct canals in the medulla and sclerites which are chiefly tuberculate clubs and sticks and spindles. This specimen thus represents the first valid record of a species of Anthothela in the southern hemisphere.
Unfortunately, defining the species is more problematic due to the assumed damage to the tips of the projecting sclerites. The cause of this partial dissolution of the sclerites is unknown. In Benham’s description there is no mention of the medium in which the specimen was originally preserved. He mentions “poor preservation” when discussing the boundary canals but does not elaborate. He also states that the “projecting points are covered with a thin stainable membrane, the mesogloea” and that the “column [calyx] is provided with short, stout, round-ended rods with many rounded knobs”. This suggests the brown, rounded, fuzzy tips on the calyx sclerites were present when Benham was originally describing the specimen and thus the damage may have already occurred. The sample was removed from an old, sealed jar for this re-description. It was found to be in 50% ethanol with trace amounts of benzene. Nevertheless, A. vickersi n. comb. can be distinguished from other species included in Anthothela by the presence of the very long, narrow sticks with sparse tubercles in the tentacle rachis and pinnules, thorn clubs in the calyx, and numerous, short, stout tuberculate rods in the cortex. This differs from A. tropicalis , which has many large and bent thorn clubs and minimal short rods in the cortex, and strongly bent, large thorn clubs in the calyx. A. quattriniae n. sp. differs by the presence of large bulbous thorn clubs. A. aldersladei n. sp. has a preponderance of short thorn clubs in the cortex and very large spindles in the points and collaret. A. pacifica and A. grandiflora do not have the knobbly surface or prickly calyces evident in A. vickersi n. comb..
Specimens of A. grandiflora carry the same haplotype as specimens of A. vickersi n. comb. using two mitochondrial gene regions (mtMutS and igr1– cox1). However, the morphological and geographical differences between these populations were deemed enough to require separation into two species. A. vickersi n. comb. has been recorded from southern Australia and New Zealand while A. grandiflora is only known from the Atlantic Ocean. The sclerites in the calyx of A. vickersi n. comb. are large, bent thorn clubs which project out from the calyx, while the sclerites in the calyx of A. grandiflora , although at times complex, do not consistently have tips which project out of the colony. Additionally, A. vickersi n. comb. has short, relatively broad, tuberculate rods common in the cortex and long, narrow sticks in the tentacles and pinnules which are not common in A. grandiflora , and the branches of the colonies of A. vickersi n. comb. are not as narrow or flexible as those in colonies of A. grandiflora .









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Anthothela vickersi ( Benham, 1928 )
| Moore, Kirrily M., Alderslade, Philip & Miller, Karen J. 2017 |
Spongioderma
| Benham 1928: 81 |