
Victorgorgia alba ( Nutting, 1908 ), 2017
|
publication ID |
https://doi.org/10.11646/zootaxa.4304.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:3D557C94-0783-4C39-80C3-9C321DA94800 |
|
DOI |
https://doi.org/10.5281/zenodo.6015385 |
|
persistent identifier |
https://treatment.plazi.org/id/3369B244-0E1F-4B6A-AB5D-9FC08F965917 |
|
taxon LSID |
lsid:zoobank.org:act:3369B244-0E1F-4B6A-AB5D-9FC08F965917 |
|
treatment provided by |
Plazi |
|
scientific name |
Victorgorgia alba ( Nutting, 1908 ) |
| status |
comb. nov. |
Victorgorgia alba ( Nutting, 1908) new combination
( Figs. 100–109 View FIGURE 100 View FIGURE 101 View FIGURE 102 View FIGURE 103 View FIGURE 104 View FIGURE 105 View FIGURE 106 View FIGURE 107 View FIGURE 108 View FIGURE 109 )
Clematissa alba Nutting, 1908: 582 View in CoL , Pl. XLIV Fig. 4 View FIGURE 4 , XLVIII Fig. 4 View FIGURE 4 . Muriceides alba ( Nutting, 1908) : Kükenthal 1924: 166.
Anthothela nuttingi nom. nov.: Bayer, 1956: 86, Figs. 9 View FIGURE 9 a–e.
NOT Anthothela alba ( Rhizoxenia alba Grieg, 1887 = Clavularia alba ): Molander 1929: 18 Material examined. Holotype: USNM 25378 About USNM , Nihoa Island , Hawaii, RV ‘ Albatross’, stn. 4157, 23.08°N, 161.87°W, depth 1394–1829 m, 6th August 1902 (“off Bird Island ” in original description). GoogleMaps
Description:
Colony form: In the original description the whole colony is described as “incomplete, about 22 mm high, consisting of a sinuous stem giving off two large unequal branches about 50 mm apart” ( Nutting 1908). The height of the colony at 22 mm would be very small and appears to be an error on Nutting’s part as the branches are then mentioned as 50 mm apart. The holotype is now two fragments, both of which are straight to slightly curved branches; one fragment has a single bifurcation ( Fig. 100 View FIGURE 100 A), the other is a small piece of branch with a terminal polyp bunch ( Fig. 100 View FIGURE 100 B). In Bayer’s redescription the colony is described as “ramose; branches stout, clavate” ( Bayer 1956a). Bayer gives the diameter of the main stem width as 6.5 mm while the branches taper from 3.5 mm in diameter proximally to 2.5 mm distally at their narrowest point below polyp clusters at the branch tip. Both authors describe the calyces as occurring on all sides of the branches (according to Nutting in an “irregular spiral”) and forming crowded clusters at the branch tip. Average space between calyces is not mentioned by either author, but they appear reasonably crowded right along the branch and occurring on all sides ( Fig. 100 View FIGURE 100 A). For the purposes of this redescription, only a tiny piece of a branch with two damaged calyces and two detached polyp heads of the holotype were available for examination ( Fig. 100 View FIGURE 100 C).
Colour: Nutting mentions the “axis, cortex and calyces are all creamy white in color (in alcohol)” but he does not mention the live colour. Bayer mentions the colour as “ivory white throughout”.
Polyps and calyces: Most of the polyps are partly retracted such that the base of the polyp head sits on the lip of the calyx ( Fig. 100 View FIGURE 100 A, B). No polyps appear to be fully retracted within the calyces and neither Nutting nor Bayer mentions such an occurrence. Nutting lists the calyx as 5.5 mm high “to the top of the operculum” and 3 mm in width at the lip of the calyx, and Bayer describes the calyces as “cylindrical and ungrooved” with no measurements. However, some calyces appear to have faint longitudinal furrows ( Fig. 100 View FIGURE 100 B). At their widest point the two polyp heads available are 3.2–3.4 mm wide and 2–2.5 mm high from the top of the calyx to the top of the polyp head. The remnant of a calyx available for this study occurs at right angles to the branch, has only a single layer of sclerites arranged in a faintly en chevron arrangement and is quite delicate ( Fig. 100 View FIGURE 100 D). In the two polyp heads available for examination and in the colony figures, the tentacles fold tightly over the polyp mouth giving the polyps a mostly flat summit with eight mounds. There are 7–9 short pinnules in a single row along each side of the tentacles.
Medulla and Cortex: The branch consists of a centrally positioned medulla, made up of tightly packed, longitudinally arranged sclerites surrounded by a cortex, also made up of a layer of packed sclerites. The cortex and medulla are clearly delineated by many, longitudinal canals forming a ring of boundary canals in cross-section ( Fig. 101 View FIGURE 101 A). These canals join and anastomose together to form a boundary space which allows the cortex to easily be separated from the medulla. Additionally, there are 4–5 large and conspicuous canals running through the centre of the medulla. These range from 0.25–0.5 mm in diameter, some are circular and some are oval. Bayer states the “solenia”, which perforate the medulla near the base, “diminish and seem to disappear entirely toward the branch tips”. No further attempt to investigate the canal arrangement was possible due to the shortage of available material.
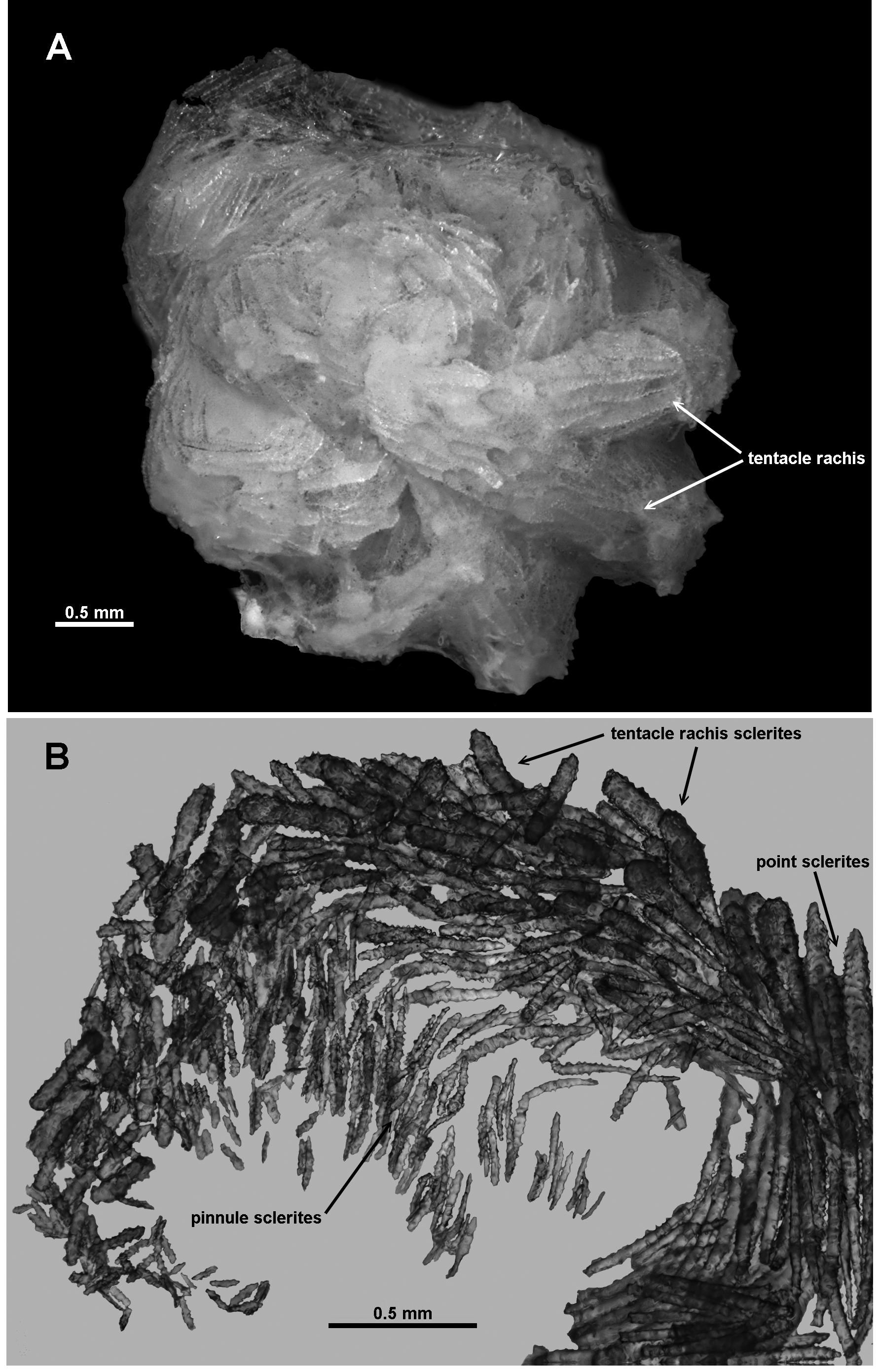
Sclerites: The polyp head is covered in a thick layer of sclerites, arranged as a collaret and points ( Fig. 101 View FIGURE 101 B). The collaret consists of approximately 8–10 transverse rows of curved to straight sticks and spindles with simple, relatively sparse tubercles ( Fig. 102 View FIGURE 102 a). Similar sclerites are arranged en chevron above the collaret, eventually becoming longitudinal at the top of the points—lengths usually range from 0.45–0.8 mm. Mixed in with these at the peak of the points are long clubs with extended tips ( Fig. 102 View FIGURE 102 b) and large, bulky clubs with reasonably crowded small warts and tubercles ( Fig. 103 View FIGURE 103 ). These are easily visible on the polyp head and make the eight points quite large and impressive ( Fig. 101 View FIGURE 101 B). The clubbed sclerites are a comparable length to the simpler sclerites in the points, usually 0.45–0.95 mm, but sclerites up to 1 mm were noted.
Sclerites are arranged longitudinally in a thick layer on the tentacle rachis and decrease in size towards the tip of the tentacle ( Fig. 104 View FIGURE 104 A, B). There are thick rods with blunt ends and crowded tubercles, as well as a few rare spindles with very few tubercles and smaller clubs with tubercles clumped at one end ( Fig. 105 View FIGURE 105 ). The thick rods are of a relatively consistent length ( 0.22–0.44mm) and appear to be more common at the proximal end of the tentacles and in the top layer of the sclerites ( Fig. 104 View FIGURE 104 B). The clubs are approximately 0.20–0.36 mm long and are more crowded in the tip and sides of the tentacles with the clubbed end arranged towards the tip of the tentacle; the mostly smooth spindles are 0.28–0.37 mm long.
Small rodlets are crowded longitudinally in the pinnules ( Figs. 104 View FIGURE 104 B; 106). Ranging from 0.09–0.26 mm long, they are lightly tuberculate, often slightly flattened, and sometimes have slightly clubbed tips with thin handles. The latter resemble josephinae clubs that are common in other species of Victorgorgia but are only poorly developed in this species ( Fig. 106 View FIGURE 106 a).
No sclerites were detected in the pharynx.
In the calyx, sclerites are straight or very slightly curved, slender sticks and spindles with only a minor covering of simple tubercles ( Fig. 107 View FIGURE 107 ). Sclerite length grades from approximately 0.32 to 0.70 mm with no noticeable size grouping on the calyx. There are also a few small, mostly smooth spindles which have a few tubercles and small thorns situated approximately mid-way on the sclerites ( Fig. 107 View FIGURE 107 a). These are shorter than the other calyx sclerites at 0.15–0.24 mm long.
Similarly, the cortex has straight, slender sticks and spindles, although these sclerites have a more substantial covering of tubercles than those from the calyx and are, in general, larger ( 0.37–0.82 mm) and thicker ( Fig. 108 View FIGURE 108 ). There are also mostly smooth spindles like those in the calyx although these are larger, ranging in length from 0.23–0.33 mm ( Fig. 108 View FIGURE 108 a).
The medulla is formed of tightly packed, longitudinally and obliquely placed sclerites that are most commonly long, straight or slightly bent sticks and spindles. The longest of these are up to 1.25 mm, but most are within 0.47– 0.98 mm ( Fig. 109 View FIGURE 109 ). Some of the sclerites are mostly smooth but with a few simple conical tubercles, and there are also spindles with more crowded warts. There is evidence of some fused and branched sclerites.
Sclerites are all transparent under transmitted light excepting the bulky sclerites in the tentacles and points, which have brown tinges.
Distribution: Hawaiian seamounts.
Depth: 1394–1829 metres.
Remarks: When Bayer (1956a) reassigned the species Clematissa alba Nutting, 1908 to Anthothela he gave it the name Anthothela nuttingi because Molander’s claim that Clavularia alba (= Rhizoxenia alba Grieg, 1887 ) belonged in Anthothela (with the resulting new combination A. alba (Grieg, 1887)) took precedence. It is now clear however, that Clavularia alba ( Grieg, 1887) does not belong in Anthothela (see below), and had Bayer known that he could have formed the binomial Anthothela alba ( Nutting, 1908) which would have been considered valid at the time. As has been demonstrated above, Nutting’s species does not belong in Anthothela and so Bayer’s reassignment of the species to that genus is no longer valid. Clematissa alba is here transferred to Victorgorgia , thus becoming V. alba ( Nutting, 1908) , and Anthothela nuttingi therefore becomes a synonym.
The holotype of Rhizoxenia alba Grieg, 1887 was examined for this study, and although no polyps remain the colony was obviously stoloniferous only, with surface sclerites quite uncharacteristic of Anthothela . Thus it is impossible to be definitive on where C. alba belongs but it appears to not be a species of Anthothela .
Unfortunately, only a tiny fragment of the holotype of V. alba n. comb. was available for examination; nevertheless, when combined with Nutting and Bayer’s descriptions it can confidently be asserted that this species does not belong in Anthothela . The presence of large canals in the medulla and the absence of sclerites in the pharynx combine to exclude this specimen from the revised definition of Anthothela . These characteristics instead support the re-assignment to Victorgorgia , as do general polyp and sclerite form. However, similar to V. macrocalyx n. comb., there are very few josephinae clubs in the tentacles, which are common in other species of Victorgorgia . This may be due to limited material or it may truly reflect that the clubs are absent and this specimen does not belong in Victorgorgia . Alternatively, it may be that the presence/ absence or abundance of the clubs is simply an interspecific variation within Victorgorgia . Specimens collected from similar locations to that of the holotype would assist in defining this quandary.
There are specimens determined as Anthothela nuttingi that have been collected around the Hawaiian Islands (S. France and A. Baco pers.com.), and some of these specimens have DNA sequences currently available on GenBank. A fragment of one such specimen ( USNM 94435 About USNM ) was examined and found not to correspond morphologically with the holotype of V. alba n. comb.. Additionally, the molecular data groups USNM 94435 About USNM with V. eminens n. sp. from the Tasmanian seamounts. Thus it is extremely likely there is more than one species of Victorgorgia on the Hawaiian seamounts and all samples previously determined as A. nuttingi require revision.
Based on what was available of the holotype, the key differences of V. alba n. comb. from other known Victorgorgia species are the presence of bulky, closely warted sclerites in the points plus thick rods on the tentacle rachis and no or very few josephinae clubs. The most comparable species is V. macrocalyx n. comb., which, unfortunately, was also restricted in material available for examination. Both species have thick rods on the tentacle rachis, bulky clubs in the points and few josephinae clubs. The differences between them include the presence of extended clubs in the points of V. alba n. comb. ( Fig. 102 View FIGURE 102 ), the tentacle rachis sclerites ( V. alba n. comb. has small clubs and bars with relatively crowded, rounded tubercles ( Fig. 105 View FIGURE 105 ) whilst V. macrocalyx n. comb. has small clubs with tall cones ( Fig. 121 View FIGURE 121 ) and squat bars with sparse tubercles ( Fig. 122 View FIGURE 122 )), and V. alba n. comb. has only slender sticks and spindles in the calyx and cortex ( Figs. 107 View FIGURE 107 ; 108) whilst V. macrocalyx n. comb. has some thicker, tuberculate spindles ( Figs. 124 View FIGURE 124 ; 125). The differences in sclerite form are sufficient to distinguish between these two small specimens here but with such limited material it has been impossible to investigate whether intracolonial variation would account for these differences.
For the other species of Victorgorgia ; V. josephinae and V. argentea n. comb. can be separated from V. alba n. comb. by their abundance of josephinae clubs and the lack of thick rods in the tentacles, V. eminens n. sp. by the absence of large, bulky sclerites in the points and V. nyahae n. sp. by the presence of sharply pointed thorn clubs.














No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
SubClass |
Octocorallia |
|
Order |
|
|
SubOrder |
Scleraxonia |
|
Family |
|
|
Genus |
Victorgorgia alba ( Nutting, 1908 )
| Moore, Kirrily M., Alderslade, Philip & Miller, Karen J. 2017 |
Clematissa alba
| Kukenthal 1924: 166 |
| Nutting 1908: 582 |