
Halocaridinides socotraensis, Fransen & Damme, 2018
|
publication ID |
https://doi.org/10.11646/zootaxa.4442.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:2C4367F2-0CD7-4559-8BBE-48948858F48D |
|
DOI |
https://doi.org/10.5281/zenodo.5957482 |
|
persistent identifier |
https://treatment.plazi.org/id/039C87DF-FFEB-FFE2-70ED-8E99FAF14B92 |
|
treatment provided by |
Plazi |
|
scientific name |
Halocaridinides socotraensis |
| status |
sp. nov. |
Halocaridinides socotraensis View in CoL sp. nov.
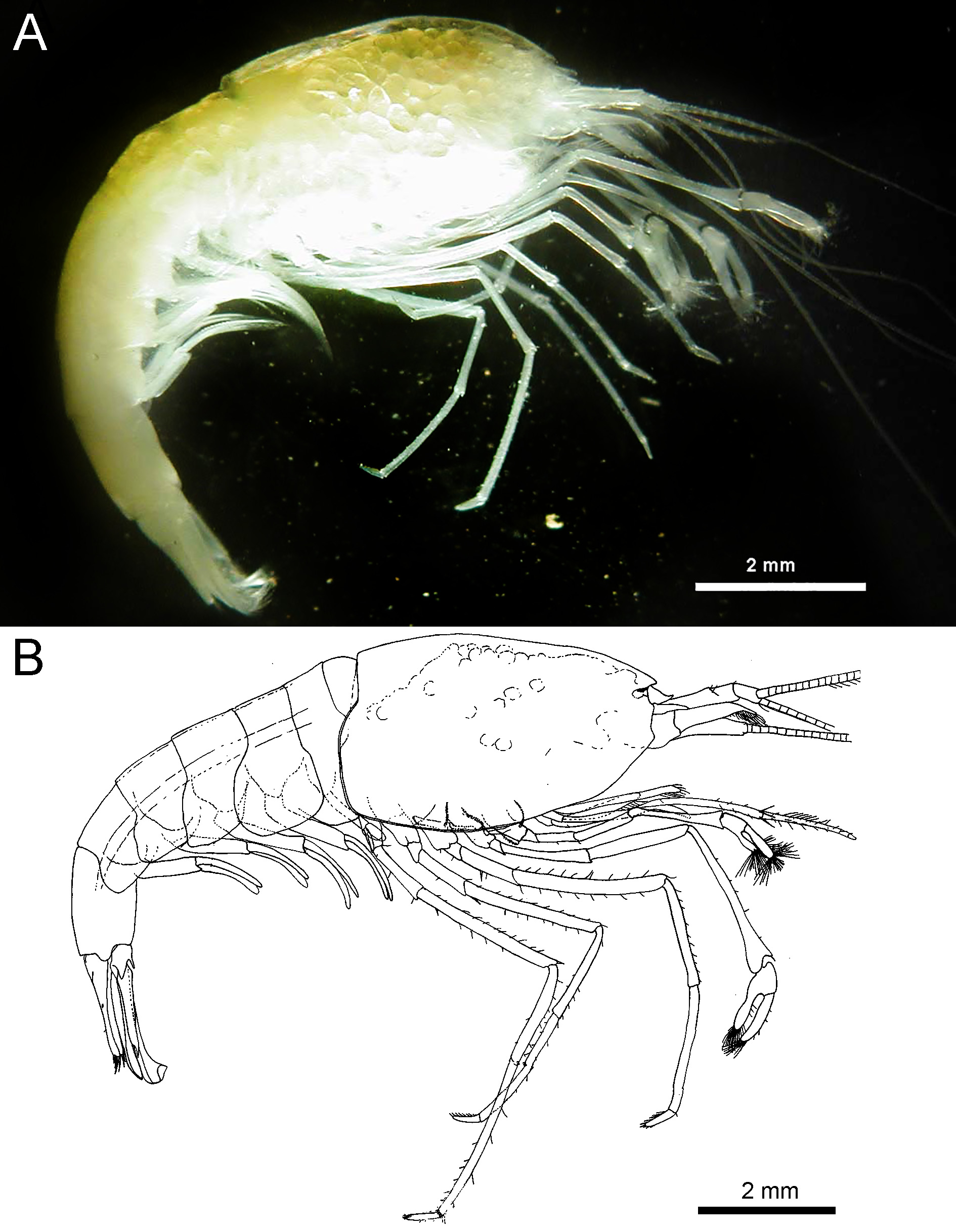
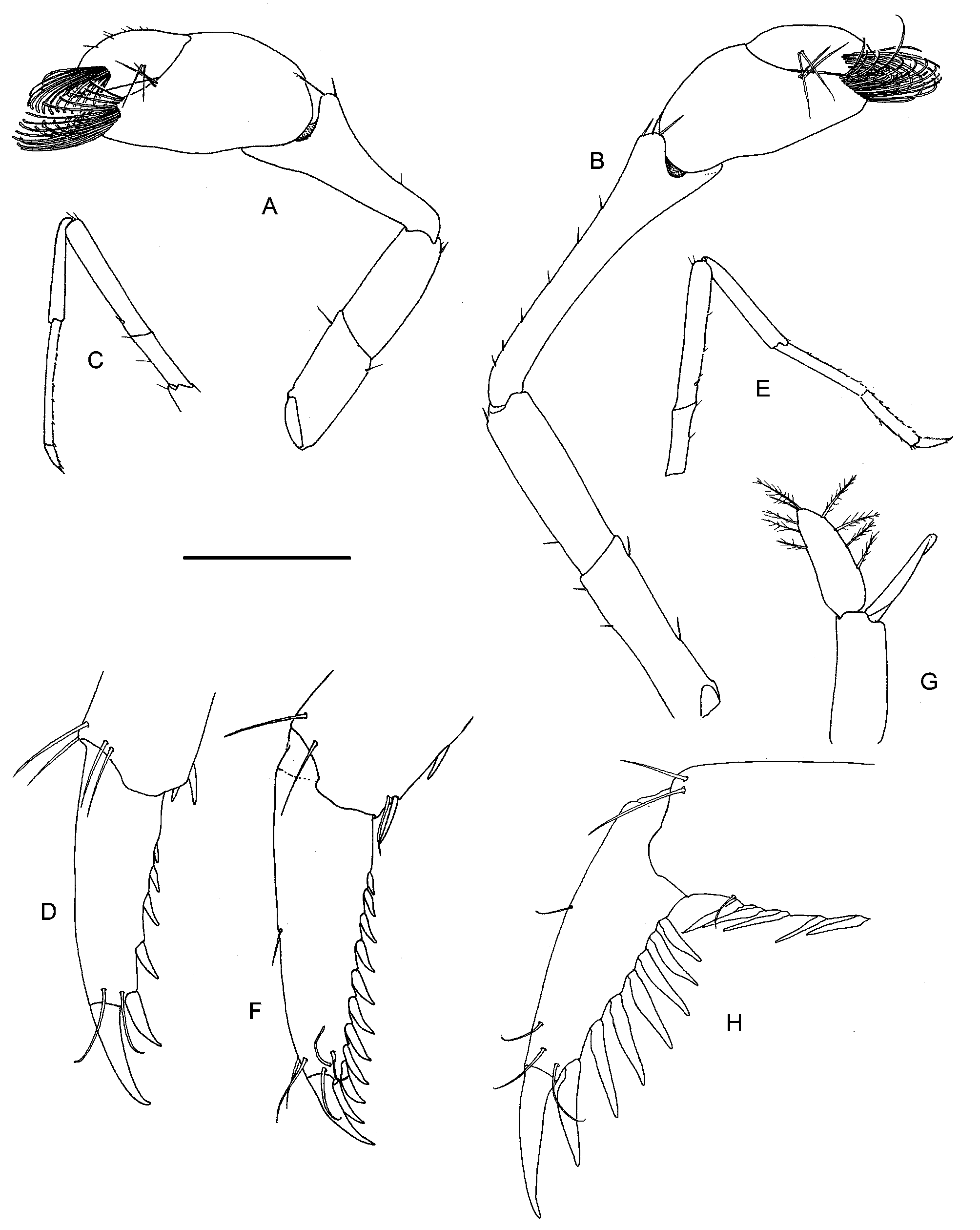
( Figs. 2–10 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 View FIGURE 9 View FIGURE 10 )
Material examined. Socotra Island ( Yemen): Ehrer Cave, 12°33’0.54”N 54°27’35.64”E, alt. 250m (type locality). 1 holotype male pocl. 3.9mm (nr. E19) RMNH GoogleMaps .CRUS.D.57274; 18 paratypes (2 males, 16 non-ovigerous females, nrs E1–18), RMNH.CRUS.D.57275: from stagnant pools near entrance, 27–28.xii.2003; collected by Kay Van Damme. 34 paratypes (24 males, 10 females of which one ovigerous, nrs E21–55), RMNH.CRUS.D.57276: pop. 1 (waterfall); 27–28.xii.2003; collected by Kay Van Damme. 6 paratypes (1 male, 5 females of which 4 ovigerous, nrs E56–59, E70, 71), RMNH.CRUS.D.57277: pop. 2 (waterfall); 27–28.xii.2003; collected by Kay Van Damme. 6 paratypes (2 males, 4 non-ovigerous females, nrs E60–65), RMNH.CRUS.D.57278: pop. 3 (endbasin); 27–28.xii.2003; collected by Kay Van Damme. Fosreher Cave, 12°37’41.1”N 53°30’42.2”E, alt. 624m, subterranean lake at - 100m. 8 paratypes (1 male, 7 non-ovigerous females, nrs F1–8) RMNH.CRUS.D.57279: - 80 m; 2.i.2004; collected by E. Claes, SKP-team. Ghiniba Cave , 12°26’01.3”N 53°56’11.5”E, alt. 395m, margin of subterranean freshwater river. 1 non-ovigerous female pocl. 3.0mm (third pereiopods missing) (nr. G1), RMNH GoogleMaps .CRUS.D.57280: 6.i.2003; collected by Kay Van Damme. 1 non-ovigerous female pocl. 4.38mm (nr. G2), RMNH. CRUS.D.57281: 9.i.2004; collected by L. Vernaeve, SKP-team. Near Qadub , 12°38’14.9”N 53°57’27.2”E, light brackish coastal well. 1 non-ovigerous female mounted in a glass slide, from a zooplankton sample, damaged (not measured), pers. coll. KVD GoogleMaps : 12.ii.1999; collected by Kay Van Damme.
Comparative material. Halocaridinides trigonophthalma ( Fujino & Shokita, 1975) . 3 specimens, RMNH . CRUS.D.40922: Japan, Akasaki-do , long narrow cave with very slowly flowing stream; width 0.6m ; 0.25m; bottom clay & stones and hard limestone rock; tourist lights intermittently on in summer; sample mainly from stagnant pool at end of cave second exit; 12x 0.6 m; 23.xii.1986; 19°C, 0 2 saturation 57%, conductivity (EC) 650 S/cm; leg. et don. N.W. Broodbakker. 2 specimens, RMNH. CRUS.D.37247: Japan, Ryukyu Islands, N coast of Yoron Island ; 25.xii.1986; leg. et don. N.W. Broodbakker. Many specimens, RMNH. CRUS.D.33133: Palau Distict , Caroline Islands, Angaur Island, Doresi Pond, 6°54’44”N 13°48’20”E; 14.vii.1980; leg. et don. J.A. Maciolek. Halocaridinides fowleri (Gordon, in Gordon & Monod, 1968) . 1 male, ZMA GoogleMaps . CRUS.D.103701: Zanzibar, from Pango Managola cave near Makunduchi ; 12.ix.1983; leg. et don. P. Beron & V. Beshkov.
Description. Body cylindrical. Carapace smooth. Rostrum ( Fig. 4A, B View FIGURE 4 ) not developed, without teeth, anterior margin of carapace rounded in dorsal view, blunt in lateral view, reaching level of rounded inferior orbital angle. Supraorbital, antennal and pterygostomian spines absent. Anterior lateral margin straight; pterygostomian angle broadly rounded ( Fig. 4A, B View FIGURE 4 ).
Abdomen smooth; pleura of abdominal segments posteroventrally rounded; sixth abdominal segment 1.5 times as long as fifth.
Telson ( Fig. 4C View FIGURE 4 ) as long as sixth abdominal segment, twice as long as its proximal width; lateral margin convex in proximal half, slightly concave in distal half; distal margin 0.6 times as wide as proximal width, straight; two pairs of dorsal spines at about 0.5 and 0.75 of telson length, about 0.0 8 times as long as telson, proximal pair submarginal, distal pair marginal; posterior margin with four pairs of spines, lateral spines as long as dorsal spines, intermediate pair long, three times as long as lateral spines, two pairs of submedian spines about twice as long as lateral spines.
Eyes ( Fig. 4A, B View FIGURE 4 ) triangular, cornea unpigmented, extending to the proximal third of basal segment of antennular peduncle.
Antennula ( Fig. 4A, B View FIGURE 4 ) with peduncle and flagella well-developed. Basal segment without distolateral tooth, distal margin not developed; medioventral tooth absent; stylocerite half as long as basal segment, with distal acute tip, slightly convex lateral margin with row of small setae, median margin sinuous, proximally rather broad; distodorsal margin of basal segment with row of short setae. Intermediate segment somewhat longer than wide, distodorsal margin with few short setae. Distal segment about as long as wide. Upper flagellum slightly longer than carapace, slender; lower flagellum about 1.3 times longer than upper flagellum, slender.
Antenna ( Fig. 4A, B View FIGURE 4 ). Basicerite of antennal peduncle with small distolateral blunt tooth; ischiocerite and mericerite normal; carpocerite cylindrical, reaching distal margin of intermediate segment of antennular peduncle; anterior lamina of scaphocerite almost reaching distal margin of distal segment of antennular peduncle, distolateral tooth distinct but small, not reaching anterior margin of lamina, lateral margin straight, anterior margin rounded.
Epistome with blunt anterior median carina; labrum oval.
Paragnath small, alae small, oblique rectangular, distomedial margin with row of setae; corpus small, unarmed.
Second to eighth thoracic sternites narrow, without special features.
Mandible ( Fig. 4D View FIGURE 4 ) with short incisor process terminating in some irregular teeth and small denticles; bristle of long setae between incisor and molar process; molar process distally concave with rows of very short setae forming transverse ridges.
Maxillula ( Fig. 4E View FIGURE 4 ) upper lacinia broad rectangular with two rows of strong spines medially; lower lacinia broadly rounded medially, with anterior lobe, with one submarginal row of short setae and several marginal rows of simple and serrate setae; palp distally bilobed with two subdistal simple setae.
Maxilla ( Fig. 5A View FIGURE 5 ) with basal endite well developed, bilobed; distal lobe much smaller than broad proximal lobe, with row of slender setae and few spines along distal and medial margin, distalmost spines distally serrate; median margin of proximal lobe with many rows of moderately long, slender, distally serrate setae; coxal endite semicircular, fringed by row of many slender setae along median margin; scaphognathite with narrow posterior lobe and broad anterior lobe, anterior lobe as long as distal lobe of basal endite; palp simple, short, subdistal seta not observed.
First maxilliped ( Fig. 5B View FIGURE 5 ) with small coxal endite with few plumose setae; basal endite long with rows of simple setae on ventral surface and row of long grid setae along median margin; caridean lobe well developed, flagellum short, both fringed by plumose setae; palp as long as caridean lobe, broad, fringed by simple setae distally.
Second maxilliped ( Fig. 5C View FIGURE 5 ) with dactylar segment triangular, median margin with long serrate setae; anterior medial margin of propodal segment produced with row of long robust setae medially; carpal segment short; meral segment with few long plumose setae medially; ischial and basal segments fused, with long plumose setae along medial margin, long plumose seta proximolaterally; exopod well developed, slightly broadened proximally with few long simple setae proximally and long plumose setae distally; coxal segment with oblong triangular epipod, without podobranch.
Third maxilliped ( Fig. 5D View FIGURE 5 ) pediform, exceeding antennal scale by ultimate segment; ultimate segment slightly shorter than penultimate segment, with one stout terminal spine and about 5 subterminal spines, with several transverse rows of strong serrate setae on medial surface; penultimate segment as long as ischiomeral segment, with row of 7 strong spines along medial margin; ischiomeral segment fused to basal segment, slightly broader than penultimate segment, with pair of long plumose setae proximomedially; exopod well developed, reaching exceeding distal margin of ischiomeral segment, with simple and plumose setae distally; coxal segment with epipod laterally and patch of plumose setae medially.
Branchial formula as follows ( Table 1):
Pereiopods. Epipods on the pereiopods are straplike grabbing the single very long robust setobranch of the next pereiopod. First pereiopod ( Fig. 6A View FIGURE 6 ) with chelae short and broad, fingers about three times as long as palm; tips of fingers with tufts of long brush-like setae; carpus 1.5 times as long as chela, distally broad, deeply excavate; merus shorter than carpus, unarmed; ischium slightly shorter than merus, unarmed; basis short; coxa with strap-like epipod and long single setobranch. Second pereiopod ( Fig. 6B View FIGURE 6 ) longer and more slender than first pereiopod; chalae as in first pereiopod; carpus twice as long as chela; merus 0.5 times as long as carpus; ischium slightly shorter than merus; basis and coxa as in first pereiopod. Third pereiopod ( Fig. 6C, D View FIGURE 6 ) with dactylus with 6 spinules along flexor margin of corpus; propodus 3 times as long as dactylus with 11 spinules along ventral margin; carpus 0.85 times propodus length, with 6 spinules along ventral margin; merus straight, slightly longer than propodus, with one spine at proximal fourth of length; ischium slightly less than half length of merus, unarmed; basis and coxa as in first pereiopod. Fourth pereiopod ( Fig. 7A, B View FIGURE 7 ) with dactylus with 7 spinules along flexor margin of corpus; propodus 3 times as long as dactylus with 11 spinules along ventral margin; carpus 0.75 times propodus length, with 5 spinules along ventral margin; merus straight, as long as propodus, with two spines at half and proximal fourth of length; ischium and basis as in first pereiopod; coxa without epipod, with setobranch. Fifth pereiopod ( Fig. 7C, D View FIGURE 7 ) with dactylus with row of 16 spinules along flexor margin of corpus; propodus with many small spines along ventral margin; propodus 4 times as long as dactylus with about 14 spinules along ventral margin; carpus 0.6 times propodus length, unarmed; merus straight, 0.8 times propodus length, with one spine in proximal fifth of length; ischium and basis as in first pereiopod; coxa without epipod nor setobranch. First pleopod ( Fig. 7E View FIGURE 7 ) with endopod as long as exopod, narrowing distally, with blunt tip, without setae. Second pleopod ( Fig. 7F View FIGURE 7 ) with appendix masculina slightly more than half as long as appendix interna, triangular, with basis broad, tapering distally, with setae along its entire median margin.
Uropods ( Fig. 4C View FIGURE 4 ) with short unarmed protopodite; exopod overreaching telson with half of distal lamina, distolateral tooth strong, only one strong mobile spine at diaeresis; endopod overreaching telson, slightly shorter than exopod.
Ovigerous females (specimens nr E21, E56–57, E67) with small clutches of 10– 15 eggs; eggs relatively large, 1.0 x 1.5mm in size. See Figs. 2–3 View FIGURE 2 View FIGURE 3 for general female morphologies.
Size. Ehrer Cave: postorbital carapace length (pocl) in largest ovigerous females is 4.06mm; pocl in largest non-ovigerous female is 5.50mm; pocl in largest male is 4.09mm; pocl of smallest male is 2.97mm; most males and females with pocl between 3 and 4 mm. Fosreher Cave: pocl of largest non-ovigerous female is 3.75mm; pocl of only male is 3.69mm. Ghiniba Cave: pocl of two non-ovigerous females respectively 4.38 and 3.06mm.
Live colouration. The specimens are a uniform milky white to beige ( Fig. 2A View FIGURE 2 ). Eggs in females white to yellow.
Etymology. The new species was named after Socotra Island ( Yemen), its type locality.
Morphological variation. The most striking variation (or cryptic speciation?) is found in the robustness of the first and second chelipeds. Differences in both the robustness of the segments of the chelipeds and of the chelae itself are present. The variation covers a relatively continuous range ( Fig. 11 View FIGURE 11 ) and the two extremes at both ends of the morphometric spectrum are: 1) tenuis type ( Figs. 2–7 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 ), specimens with slender P1 and P2 and with chela with the fingers 2–3 times the palm length, and the chela 3-5 times longer than the palm height, the fingers are usually gaping, the carpus is long and relatively narrow distally. Bristles on the propodus are relatively long, the longest about two thirds of the propodus length ( Fig. 6A View FIGURE 6 ); 2) robustus type ( Figs. 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10 ), robust chela with fingers about as long as the palm, and the chela 2.0–2.5 times the palm height, the fingers are not gaping, the carpus is short and relatively broad distally. Bristles on the propodus are relatively shorter, up to half the propodus length (fig. 10A). These two types have no taxonomical value at this point.
There has been no correlation found of these types with either sex or size of the specimens ( Table 2; Fig. 11 View FIGURE 11 ). Animals with both slender and robust chelae were found in Erher Cave in stagnant pool and waterfall localities, as well as in Fosreher Cave and Ghiniba Cave. Other features are rather constant. The number of mobile spines at the uropodal diaeresis is always one. The position of the proximal dorsal pair of spines on the telson is between 0.5 and 0.6 of the telson length, that of the distal pair 0.7–0.8. Only few specimens have the distal pair somewhat closer to the distal margin ( Fig. 9A View FIGURE 9 ).
All female specimens have short forward-directing spinules placed on the flexor margins of the dactyli of the third, fourth and fifth pereiopods and short spines on the distoventral margin of the propodi ( Fig. 10D, F View FIGURE 10 ). Most males (n=20) have long spinules placed perpendicular on the corpus and long spines on the distoventral margin of the propodi as well. The other males (eight) have these features as in the females. These morphological differences in males are not correlated with size nor with location. The length of the appendix masculina ranges from 0.5–1.5 times the length of the appendix interna. There is no relation between the size of the appendix masculina with that of the appendix interna and the variation in dactylar morphology in males.
......continued on the next page
Distribution. Socotra Island, Yemen. Found in four localities ( Fig. 1 View FIGURE 1 ): northwest (Fosreher Cave in the Maleh or Ma’alah Plateau), north central (coastal well near Qadub), northeast (Erher Cave in the northern escarpments of the Momi Plateau) and south central (Ghiniba Cave in the Shibehon Plateau). Most common in Erher Cave.
Ecology. Subterranean freshwater and slight brackish environments in limestone karst. In Erher, populations were found in three separate locations in the cave: large densities were found in temporary shallow stagnant muddy pools near the entrance that were created by overflow during the rainy seasons. The pools near the entrance receive some light from the cave entrance, yet animals remained unpigmented, and bright white. Lower densities in the clear, shallow permanent water were found in Erher Cave at the base of the large waterfall (creating a strong flow; this can be seen as riverine) and only few animals in the clear, deep permanent lake (end basin) at the end of the cave (no flow). The water in Erher derives from the Momi plateau above and is fresh, measuring between 400–600 µS/cm (256–384 ppm) and a pH of 7.6–7.8. The waterfall is used as local water source ( De Geest & Van Damme 2006), transporting water through tubes to the nearby village (Irisseyl). In Ghiniba Cave only two specimens were found in flowing water in the large and clear permanent cave river at the end of the dry section of the cave in the main gallery. The river in Ghiniba contains freshwater of the same low conductivity as in Erher. Conductivity of the water in Fosreher Cave was not measured, yet this can be considered as freshwater as well and was used as drinking water by the team (not brackish in taste). Only few animals (eight) were found in the latter locality, which like Ghiniba functions as a sump for the runoff from the surrounding plateau. Water in the coastal well (near Qadub) was slightly brackish (4200 µS/cm, corresponding to ca. 2690 ppm) and near neutral pH (7.5), here only one specimen was found. Reproductive biology not studied, ovigerous females have large egg clutches containing many white to bright yellow eggs.
Remarks. The new species differs from H. trigonophthalma in having only one spine at the uropodal diaeresis in all specimens while H. trigonophthalma has 1–9 spines ( Fujino & Shokita 1975, Naruse et al. 2003, present material); in the strongly reduced rostrum, in the absence of pigmentation in the eye; in the appendix masculina becoming not longer than 1.5 times the length of the appendix interna while more than three times as long in H. trigonophthalma (present material). H. socotraensis sp. nov. differs from H. fowleri in having only one spine at the uropodal diaeresis in all specimens while H. fowleri has a variable number of 1–3 spines there. In H. fowleri the rostrum is acute in lateral aspect whereas it is blunt in the new species. The endopod of the first pleopod in males of the new species is almost as long as the exopod and devoid of marginal setae while in the only male specimen known of H. fowleri it is half as long as the exopod bearing several setae along the median margin.
The recently described genus from Madagascar, Monsamnis Richard, De Grave & Clark, 2012 also has only a single spine in the uropodal diaeresis ( Richard et al. 2012), like the new species of Halocaridinides from Socotra. However, in Monsamnis all pereipods have exopods (fifth reduced), whereas in Halocaridinides exopods are absent on all pereiopods. Monsamnis is known from freshwater mountain streams on Madagascar.









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
InfraOrder |
Caridea |
|
Family |
|
|
Genus |