
Chilocoris monticola, Imura, Jimpei, 2011
|
publication ID |
https://doi.org/ 10.5281/zenodo.206103 |
|
DOI |
https://doi.org/10.5281/zenodo.6182566 |
|
persistent identifier |
https://treatment.plazi.org/id/039CCB09-FFE9-9D20-439A-727C97DAEEFC |
|
treatment provided by |
Plazi |
|
scientific name |
Chilocoris monticola |
| status |
sp. nov. |
Chilocoris monticola , new species
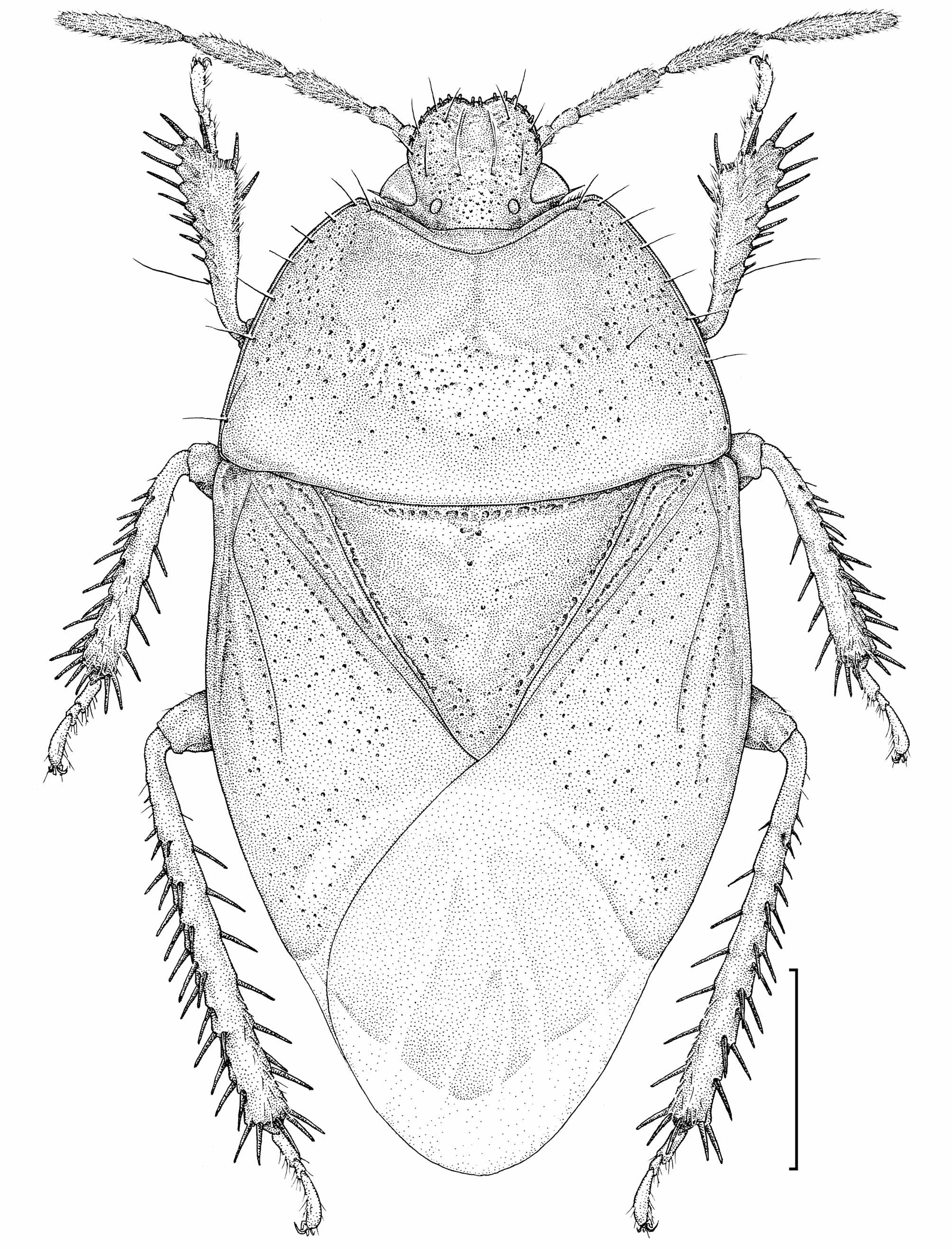
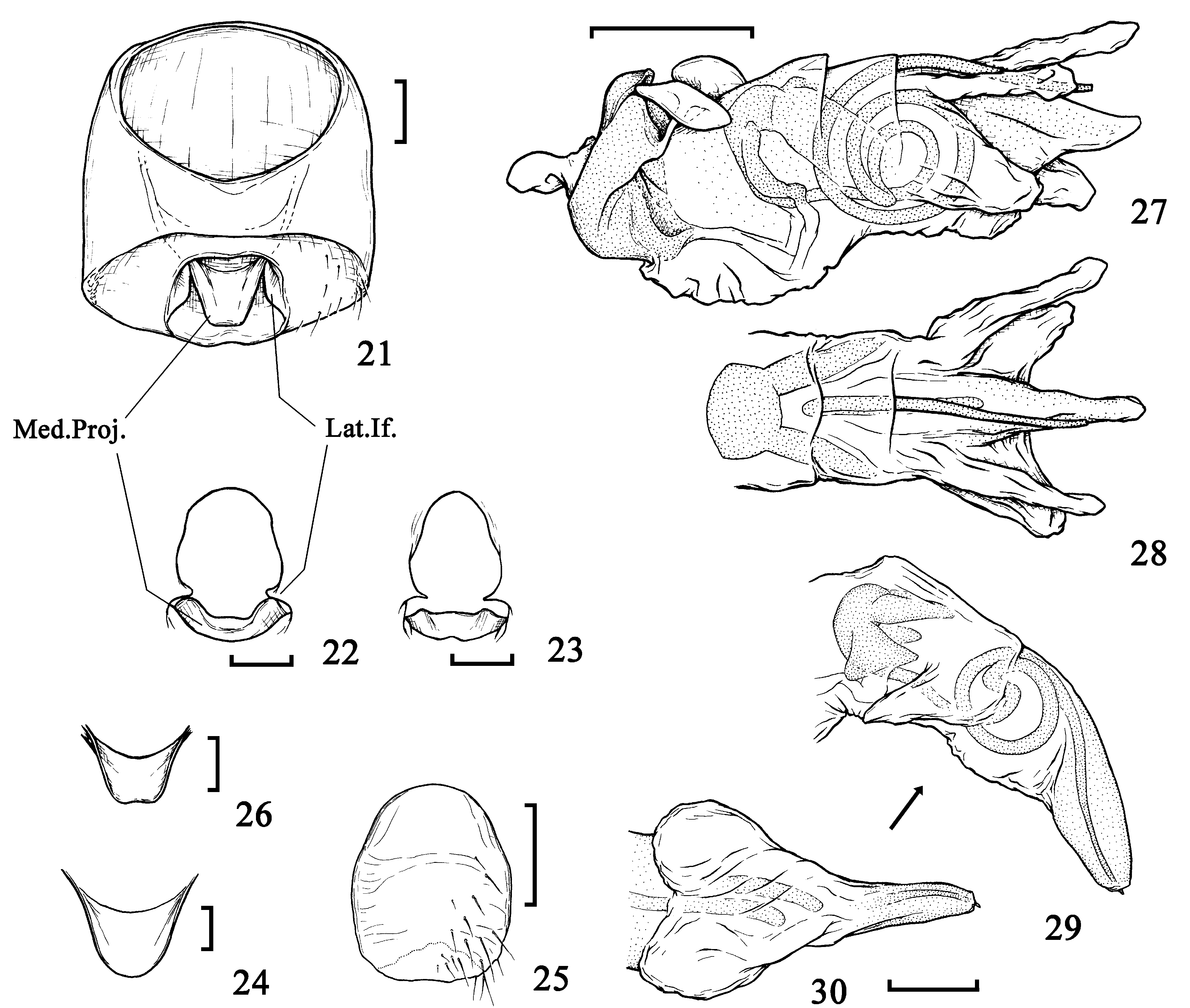
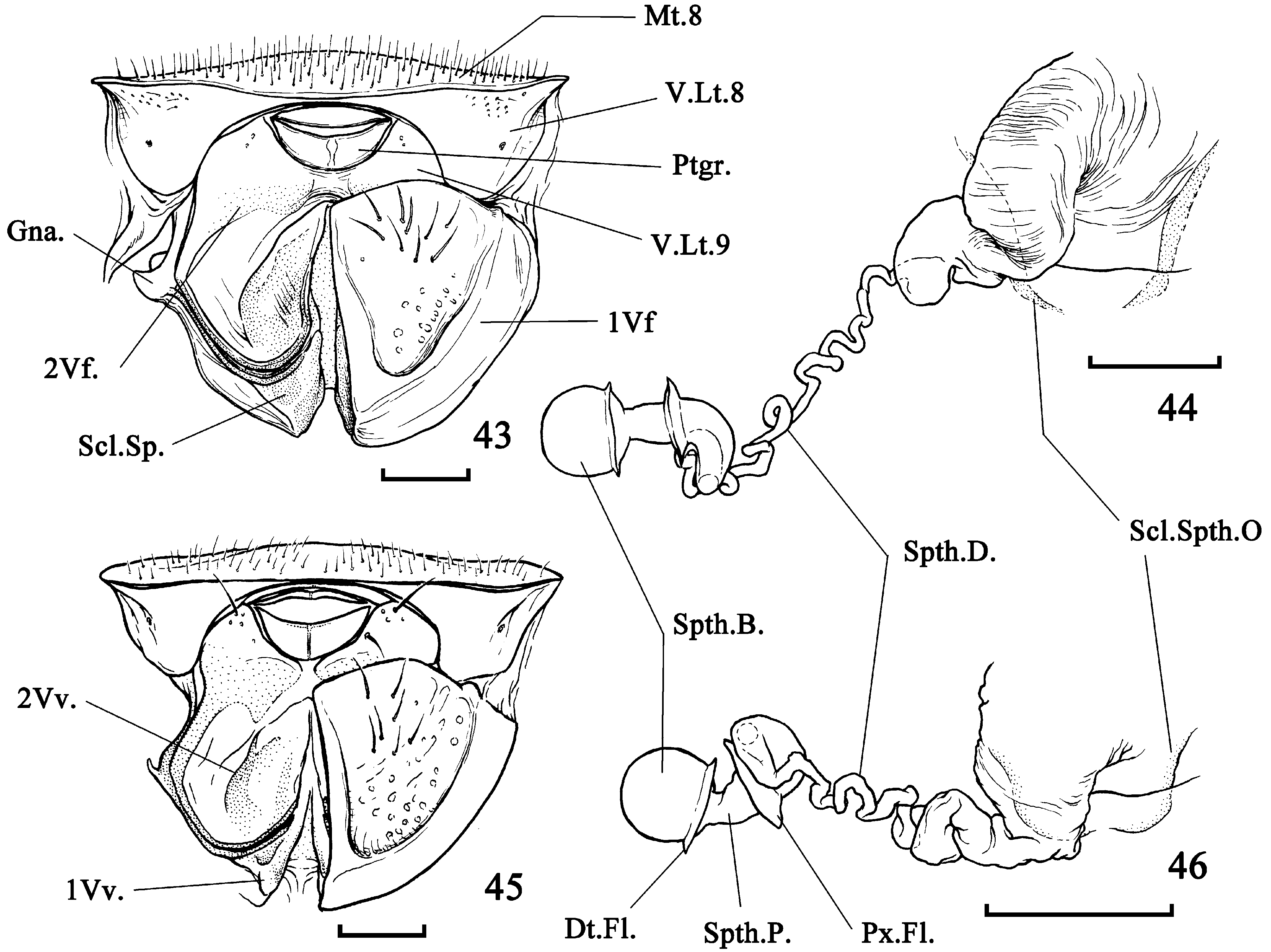
( Figs. 6–8 View FIGURES 1 – 10 , 17–22 View FIGURES 15 – 20 View FIGURES 21 – 30 , 37–39 View FIGURES 31 – 42 , 45–47 View FIGURES 43 – 46 View FIGURE 47 )
Chilocoris nitidus auct . (recorded from Japan), non Mayr.
Diagnosis. Total length 4.50–5.50. Arrangement of paraclypeal secondary setae, in order from mesalmost to lateralmost: 3 pegs - 1 hair - 2 pegs - 1 hair - 1 peg ( Fig. 6 View FIGURES 1 – 10 ). Pronotum ca. 1.8× as wide as long, tapered anterad, with 6–7 setae on each lateral submargin ( Fig. 48 View FIGURES 48 – 49 ). Hind wing without Icl ( Fig. 7 View FIGURES 1 – 10 ). 3: Phallus: endphallic duct long, protruding from dorsum of endosoma ( Fig. 17 View FIGURES 15 – 20 ); conjunctival appendage 2-lobed ( Fig. 20 View FIGURES 15 – 20 ). Paramere ( Fig. 37–39 View FIGURES 31 – 42 ) claw-like in distal part. Ƥ: 2nd valvifer angular in lower side ( Fig. 45 View FIGURES 43 – 46 ). Spermathecal duct shorter than that of C. birmanus ( Fig. 46 View FIGURES 43 – 46 ).
This species should be allied with C. nitidulus and C. nitidus . However, it can be distinguished from C. nitidulus by the male genital characters: namely, the length of endophallic duct, the arrangement of conjunctival appendages, and the shape of paramere (see also description of C. nitidulus , below). On the other hand, it differs from C. nitidus in the shape of paramere (see Lis, 1994: Fig. 124). In addition, Lis & Heyna (2001: Fig. 43 View FIGURES 43 – 46 ) reported the hind wing of C. nitidus to possess two interclaval veins, which are absent in C. monticola ( Fig. 7 View FIGURES 1 – 10 ).
Description. Holotype 3—Total length 5.50; total width 2.80; body 5.10 long, 1.50 high; head 0.80 long, 1.08 wide; antennal segments I 0.30, II 0.17, III 0.45, IV 0.48, V 0.60; rostral segments I 0.38, II 0.63, III 0.55, IV 0.40; pronotum 1.50 long, 2.65 wide; scutellum 1.30 long, 1.55 wide.
Body dark brown to blackish; posterior border of pronotum and corial area more brownish. Eyes reddish; ocelli yellowish. Antennae more brownish in segments I, II and V. Rostrum dark brown. Tarsi brown. Hemelytral membrane hyaline, obviously tinged with brown.
Body ( Fig. 47 View FIGURE 47 ) elliptical, ca. 1.9× as long as wide; punctures on dorsum small and fine, similar in size. Head ( Fig. 6 View FIGURES 1 – 10 ) dorsally punctured; clypeus slightly shorter than paraclypei; arrangement of paraclypeal secondary setae, in order from mesalmost to lateralmost: 3 pegs - 1 hair - 2 pegs - 1 hair - 1 peg. Ocular index 2.9; ocellar index 6.1; interocellar index 4.2. Venter of head almost smooth; buccula moderately developed, surpassing rostral segment I posteriorly, and almost as high as rostral segment II; maxillary plate weakly striate near buccula, clearly separated from paraclypeus with distinct suture; paraclypeal venter flattish, covered with minute hairs. Antennal segment III 2.7× as long as II. Rostrum surpassing middle of mesosternum. Pronotum ca. 1.8× as wide as long, tapered anterad, bearing 7 setae on lateral submargin, and 1 seta on lateral part of anterior submarginal line, in each side ( Fig. 47 View FIGURE 47 ). Propleuron almost smooth, but with some striae in depression. Prosternum coarse; prosternal carinae distinct. Scutellum moderately covered with punctures; punctuation weaker in medio-basal part ( Fig. 47 View FIGURE 47 ). Hemelytron well surpassing abdominal tip; punctures on corial area forming rows along either side of R, Cu and 1A, and covering remaining part of mesocorium ( Fig. 47 View FIGURE 47 ); Cu depressed in proximal part. Meso- and metapleura ( Fig. 8 View FIGURES 1 – 10 ) almost smooth; anterior margin of mesopleural evaporatorium more or less concave in lateral part; peritreme moderately recurved at apical 1/3. Meso- and metasterna sparsely covered with thin hairs; mesosternum convex. Legs slender; femur covered with hairs, without spines. Abdomen: sterna III–VII smooth, sparsely covered with minute hairs. Pygophore: ventro-posterior margin slightly producing medially.
Paratypes Ƥ —Total length 4.50–5.50; total width 2.40–3.00. Similar to holotype. Body 1.8–1.9× as long as wide. Ocular index 2.9–3.4. Antennal segment III 2.25–2.90× as long as III. Hind wing without Icl ( Fig. 7 View FIGURES 1 – 10 ).
3 terminalia—Pygophore ( Fig. 21 View FIGURES 21 – 30 ): posterior surface sparsely to moderately covered with thin hairs on lateral part; ventro-posterior margin weakly reflex, sometimes weakly producing medially; lateral infoldings of opening extended toward pygophoral lumen above parameral socket; median projection somewhat narrowed, truncate or rounded apically; opening broader than that of C. birmanus ( Fig. 22 View FIGURES 21 – 30 ). Phallus ( Figs. 17–19 View FIGURES 15 – 20 ): endosoma tapered and peaked apically, with sclerotized band distal to conjunctival appendage; this band upcurved apically ( Fig. 19 View FIGURES 15 – 20 ); conjunctival appendage 2-lobed; its dorsal lobe totally membranous, and ventral one moderately sclerotized ( Fig. 20 View FIGURES 15 – 20 ); endophallic duct long, coiled 4 times from its turning point, and protruding from dorsum of endosoma ( Fig. 17, 19 View FIGURES 15 – 20 ). Paramere ( Figs. 37–39 View FIGURES 31 – 42 ) slender; mesal margin roughly straight through the way to hypophysis in ventral view; hypophysis sharp, claw-like, directed dorsally. Proctiger broader than that of C. birmanus .
Ƥ terminalia ( Fig. 45 View FIGURES 43 – 46 )— Mediotergite VIII covered with thin hairs; ventral laterotergites VIII narrowly connected with each other above succeeding segments, covered with small punctures near dorsal margin; spiracle VIII reduced, but visible. 1st valvifer bearing several setae and covered with punctures in posterior surface; mesal margin elevated. 1st valvula possessing well developed inner and outer rami, with obviously pigmented spot in angle between these rami; triangulin broad, smooth, slightly sclerotized. Ventral laterotergite IX and 2nd valvifer continuously fused; this unit angular in lower side, extended mesad and approaching that of another side beneath proctiger; gonangulum thin, curved. 2nd valvula with well sclerotized ramus; several thin hairs arranged alongside the ramus on apical part. Proctiger: sternum totally split along midline. Ring sclerites well separated, lying near lateral ends of genital chamber. Spermatheca ( Fig. 46 View FIGURES 43 – 46 ): duct crinkly, divided between proximal thicker portion and distal thinner one; distal flange present on basal part of bulb; bulb spherical.
Type series. Holotype 3: “[Honsyû[!], JAPAN] / Sakai, Fujimi-machi, / Suwa-gun, Nagano Pref. / 14~ 15.VIII.2007 / Jyunnosuke KANTOH leg.” ( Figs. 6, 8 View FIGURES 1 – 10 , 47 View FIGURE 47 ) (TUA).
Paratypes 24333Ƥ—JAPAN: Honshû: [Aichi Pref.] Tsugu, Shitara: 13, 20.vii.2008, M. Yazaki leg. (TUA). [Gumma Pref.] Hanasaku, Katashina-mura: 1Ƥ, 9.vii.2009., T. Ban leg. (TUA); 536Ƥ (232Ƥ coupled), 2.viii.2007, J. Imura leg. (1Ƥ, Fig. 7 View FIGURES 1 – 10 ; 13, Figs. 17–22 View FIGURES 15 – 20 View FIGURES 21 – 30 ) (TUA); Ikaho: 13, 24.vii.1954, S. Katsuya leg. (TUA); Mt. Haruna-san: 1Ƥ, 25.vii.1954, S. Katsuya leg. (TUA); nr. Nidoage: 13, 13.viii.1962, Y. Watanabe leg. (TUA); Yubiso, Minakami-machi: 1Ƥ, 8.vi.2009., J. Imura leg. (TUA). [Ibaraki Pref.] “UWADAI-nursery”: 13, 23.vii.1952, at light (TUA). [Kanagawa Pref.] Fudakake, Katashina-mura: 1Ƥ, 8.vii.2006., J. Imura leg. (TUA); Uenohara: 13, 19.vi.1963, S. Katsuya leg. (TUA). [Nagano Pref.] Kamikochi, Matsumoto-shi: 13, 23.vii.1951., T. Nakane leg., det. as Chilocoris nitidus Mayr by H. Hasegawa ( NIAES). [Nara Pref.] Dorogawa, Tenkawa-mura: 1Ƥ, 12.viii.1958., I. Hiura leg. ( NIAES). [Nara–Mie Pref.] Mt. Ôdaigahara: 1Ƥ, 1.viii.1953., O. Sato leg. ( NIAES). [Saitama Pref.] Hashidate, Chichibu-shi: 1Ƥ, 17.vii.1999, S. Arai leg. (TUA); Onouchi-zawa-keikoku, Onouchi: 1Ƥ, 27.vii.2007., T. Iwata leg. (TUA); Ôyamabuchi, Ôtaki-mura, Saitama Pref.: 13, 31.vii.1999., S. Arai leg. (TUA). [Shizuoka Pref.] Yugashima, Izu-shi: 233Ƥ, 3.vi.2007., J. Imura leg. (TUA). [Tochigi Pref.] Nikko: 1Ƥ, 13.viii.1961, S. Tachikawa leg. (TUA). [Tokyo Pref.] Mt. Takao-san: 1Ƥ, 12.vii.1961. K. Mizusawa leg. (TUA); 13, 17.vi.1952., N. Fukuhara leg. ( NIAES). [Wakayama Pref.] Mt. Koya-san: 1Ƥ, 3.viii.1958., I. Hiura leg. ( NIAES); 1Ƥ, 26.viii.1955., S. Gotoh leg. ( NIAES). [Yamagata Pref.] nr. Atsumi: 13, 27.vii.1960, Y. Watanabe leg. (TUA). [Yamanashi Pref.] Fuji-rindou, Narusawa-mura: 13, 9.vii.2009., H. Katahira leg. (TUA); Kanayamadaira, Hokuto-shi: 133Ƥ, 7. viii. 2008., S. Fujinuma leg. (TUA); Nakayama Pass, Hokuto-shi: 135Ƥ, 7.vii.2007., J. Imura leg. (1Ƥ, Fig. 45–46 View FIGURES 43 – 46 ) (TUA). Shikoku: [Ehime Pref.] Ishizuchi Skyline, Kumakôgen-chô: 131Ƥ, 9.vii.2006., S. Yano leg., by truck mounted trap 17:30–18:30 (OMM); Jôju-sha: 1Ƥ, 25.vii.1963, M. Yasukawa leg. (TUA); Kanmon, Omogo-mura: 131Ƥ, 17.vii.2002., S. Yano leg., by light trap (OMM); Tochiwara, Omogo-mura: 13, 3.viii.2000., S. Yano leg. (OMM). [Tokushima Pref.] Mt. Tsurugi-san: 1Ƥ, 30.vii.1953., Y. Nishioka leg. ( NIAES). Kyûshû: [Fukuoka Pref.] Mt. Hiko-san, Soeda-machi: 13, 8–11.ix. 1997., T. Ishikawa leg. (TUA). [Miyazaki Pref.] Tabaru, Takachiho-chô: 13, 21–23.vi. 2008., T. Ishizaki leg. by flight interception trap (TUA).
Etymology. It is named for its occurrence in a montane habitat.
Distribution. Japan (Honshû, Shikoku, Kyûshû).
Remarks. Until recently, this species has been recognized as C. nitidus in Japan, even as Lis (1994, 1999a, 2006b) repeatedly mentioned doubts about its identity. It was first reported from Japan by Yamashita et al. (1972) on the basis of a specimen identified by H. Hasegawa as C. nitidus . Unfortunately, this exact specimen could not be examined. Other specimens, also identified as C. nitidus by H. Hasegawa, were examined instead, leading to conclusions that these represent an undescribed species. Incidentally, C. nitidus was also recorded from Japan by Distant in 1883, and he again referred to the record in 1902. However, the specimen he examined is probably C. confusus Horváth , another Japanese representative of the genus (see Horváth 1919: p. 262).
Biology. As can be inferred from the specific name, this species inhabits mountainous regions. Type materials were frequently found in moderately wet soil on the forest floor, in altitudes of more than 500 m. Specimens were sometimes attracted to artificial light.
Scant information is available to construct its lifecycle, but it is probable that emergence occurs from late May to early June at least in Central Japan, because fifth instars have been collected on May 22 from Yamanashi Prefecture, and teneral materials on June 3 from Shizuoka Prefecture. Adults appear to be active during July to August; coupling individuals have been observed on August 2 in Gunma Prefecture.




| NIAES |
National Institute for Agro-Environmental Sciences |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |