
Ellobius, FISCHER, 1814
|
publication ID |
https://doi.org/ 10.26879/1122 |
|
persistent identifier |
https://treatment.plazi.org/id/039CFE7F-0875-DA28-FF3F-FD9B7ACA0FFF |
|
treatment provided by |
Felipe |
|
scientific name |
Ellobius |
| status |
|
THE GENUS ELLOBIUS FISCHER, 1814 View in CoL
Distribution and Ecological Remarks of the Extant Ellobius Species
Nowadays, the genus Ellobius Fischer, 1814 , occurs in southeast Europe, western and central Asia with five species ( Coşkun, 2001, 2016; Wilson et al., 2017, Kaya et al., 2018): E. talpinus ( Pallas, 1770) , E. tancrei Blasius, 1884, E. alaicus Vorontsov et al., 1969, E. fuscocapillus ( Blyth, 1843) and E. lutescens Thomas, 1897 . These fossorial species inhabit steppes, grasslands and semi-deserts, and are highly adapted to subterranean life (Kryštufek and Vohralík, 2009; Coşkun, 2016).
In Iran, where the Kaldar Cave is located and the fossil material under study come from, Ellobius is currently represented by E. lutescens , E. fuscocapillus and E. talpinus ( Gharkheloo, 2003; Firouz, 2005; Kryštufek and Vohralík, 2009; Kryštufek and Shenbrot, 2016; Rusin, 2017).
Ellobius lutescens (western mole vole) is distributed in northwestern Iran, Iraq, Azerbaijan, Armenia and eastern Anatolia ( Thomas, 1905; Ellerman and Morrison-Scott, 1951; Darlington, 1957; Osborn, 1962; Walker, 1964; Lay, 1967; Hassinger, 1973; Roberts, 1977; Corbet, 1978; Corbet and Hill, 1991; Coşkun, 1997; Nowak, 1999; Wilson and Reeder, 2005; Kryštufek and Shenbrot, 2016; Wilson et al., 2017). In Iran, this species is found in mountain grasslands, sandy semi-deserts and steppe areas ( Kryštufek and Shenbrot, 2016; Tesakov, 2016).
Ellobius fuscocapillus (southern mole vole) shows a range across northeastern Iran, Turkmenistan, Afghanistan and Pakistan. In Iran it is found in open steppes with loose soil ( Gharkheloo, 2003; Shenbrot et al., 2016).
Ellobius talpinus (northern mole vole) is distributed in southeastern Ukraine and Russia, Kazakhstan, Uzbekistan, Turkmenistan and in the small part of northern Iran. Its habitat requirements are similar to that of Ellobius lutescens ( Rusin, 2017) .
The geographical areas occupied by the three species show differences in mean annual temperatures and precipitations ( Table 1). Ellobius talpinus is found in regions with drier conditions and lower maximum annual temperature than that of the other two species. The geographic ranges of E. fuscocapillus and E. lutescens display similar temperatures, but E. fuscocapillus occurs in wetter environments. All the temperatures and precipitation levels are estimations, consistent with the above-described type of habitat. However, since their subterranean life makes them relatively insensitive to high variations in surface temperatures and precipitations, we can consider that all three species have essentially the same habitat requirements. What can make differences is the resulting vegetation cover, which is of course important for the survival of the animals. But all what we can assume in the current state of knowledge is that the Ellobius species provide significant information as indicators for steppe environments. Further studies are needed to evaluate more precisely the potential of the different Ellobius species as palaeoenvironmental and palaeoclimatic indicators.
Fossil Record of Ellobius
Arvicolines are commonly used in Quaternary biostratigraphy because of their rapid evolution and their abundance in the fossil record. The genus Ellobius may represent a crucial biostratigraphic yardstick in the Zagros mountain range, which marks the western limits of its extant distribution range in the western Asia ( Weissbrod and Weinstein-Evron, 2020). However, the palaeobiogeographic and stratigraphic range of the genus in this region is still debated.
Remains of cf. Ellobius have been reported in early Pliocene (Ruscinian) sites in Kotovka, Odessa in the Ukraine ( Nesin and Nadachowski, 2001), and in two late Pliocene (Late Villanyian) sites, in the west of Ukraine and in Rivoli Veronese, northeastern Italy ( Sala et al., 1994). However, Tesakov (1998) believes that the Ellobius record from Italy belongs to Ungaromys dehmi .
According to several authors (compiled in Topachevsky and Rekovets, 1982; Maul and Markova, 2007; Tesakov, 2016), there was a succession of several Ellobius species during the early Pleistocene in Eastern Europe: E. paleotalpinus, E. melitopoliensis, E. primigenis, E. lakhutensis, E. tauricus, E. kujalnikensis and E. tarchancutensis. The morphology of E. tarchancutensis suggests that it could be the ancestor of E. lutescens (Topachevsky and Rekovets, 1982; Tesakov, 2016).
In western Asia, Ellobius has been recognized in several Middle Pleistocene archaeological sites in Sel’-Ungur in Kyrgyzstan ( Ellobius ex gr. tancrei; Markova, 1992), in Hummal layer G in Syria ( Ellobius sp. ; Maul et al., 2015), Azokh-1 units Vm, Vu, III, II/III, II ( Ellobius sp. ; Fernández-Jalvo, 2016) in Nagorno Karabakh and Krasarin ( Ellobius (Bramus) pomeli ; Tesakov, 2016).
During the Middle Pleistocene, Ellobius extended its range westwards to Israel, Tabun C and D ( Bate, 1937; Frumkin and Comay, in press), Misliya Cave ( Weissbrod and Weinstein-Evron, 2020) and North Africa as far as to the Moroccan Atlantic coast ( Jaeger, 1988; Stoetzel, 2013). It probably arrived there from western Asia via the Libyco-Egyptian route during a cooling and aridification of North Africa favouring the development of steppes at the beginning of the Middle Pleistocene ( Stoetzel, 2013). The North African species then evolved independently from the Asian ones, through the succession of E. africanus, E. atlanticus, E. barbarus and E. zimae ( Jaeger, 1988). In most studies, it is concluded that Ellobius , outside its current range, disappeared at the end of the Middle Pleistocene at the latest ( Stoetzel, 2013; Maul et al., 2015; Weissbrod and Weinstein-Evron, 2020).
In the Late Pleistocene, Ellobius trancrei has been described from the Mousterian site of Ogzy- Kichik, Tadzhikistan ( Markova, 1992). The modern species E. lutescens was found in Iraq ( Bate, 1930) in the layers of Hazar Merd, dated to 25,000 years ago ( Coşkun, 2016). Hashemi et al. (2006) noted that remains of E. lutescens have been found in several Late Pleistocene and early Holocene sites in western and northwestern Iran: Kani Mikaeil (Kordestan), Qalaloun near Kouhdasht, Yafteh Gar, and Arjeneh near Khoramabad (Lorestan). Ellobius lutescens has also been documented in Upper Palaeolithic and Neolithic units of Dzudzuana Cave ( Georgia, 34.4-6 ka cal BP; Belmaker et al., 2016), and also in Aghitu-3 level VII ( Armenia, Upper Palaeolithic, 39-36 ka cal BP; Kandel et al., 2017). Ellobius sp. has been described in Azokh-1 unit I (157 ± 26 ka BP) and in the Holocene site Azokh-5 (Nagorno Karabakh, Parfitt, 2016).
Description of Tooth Morphology
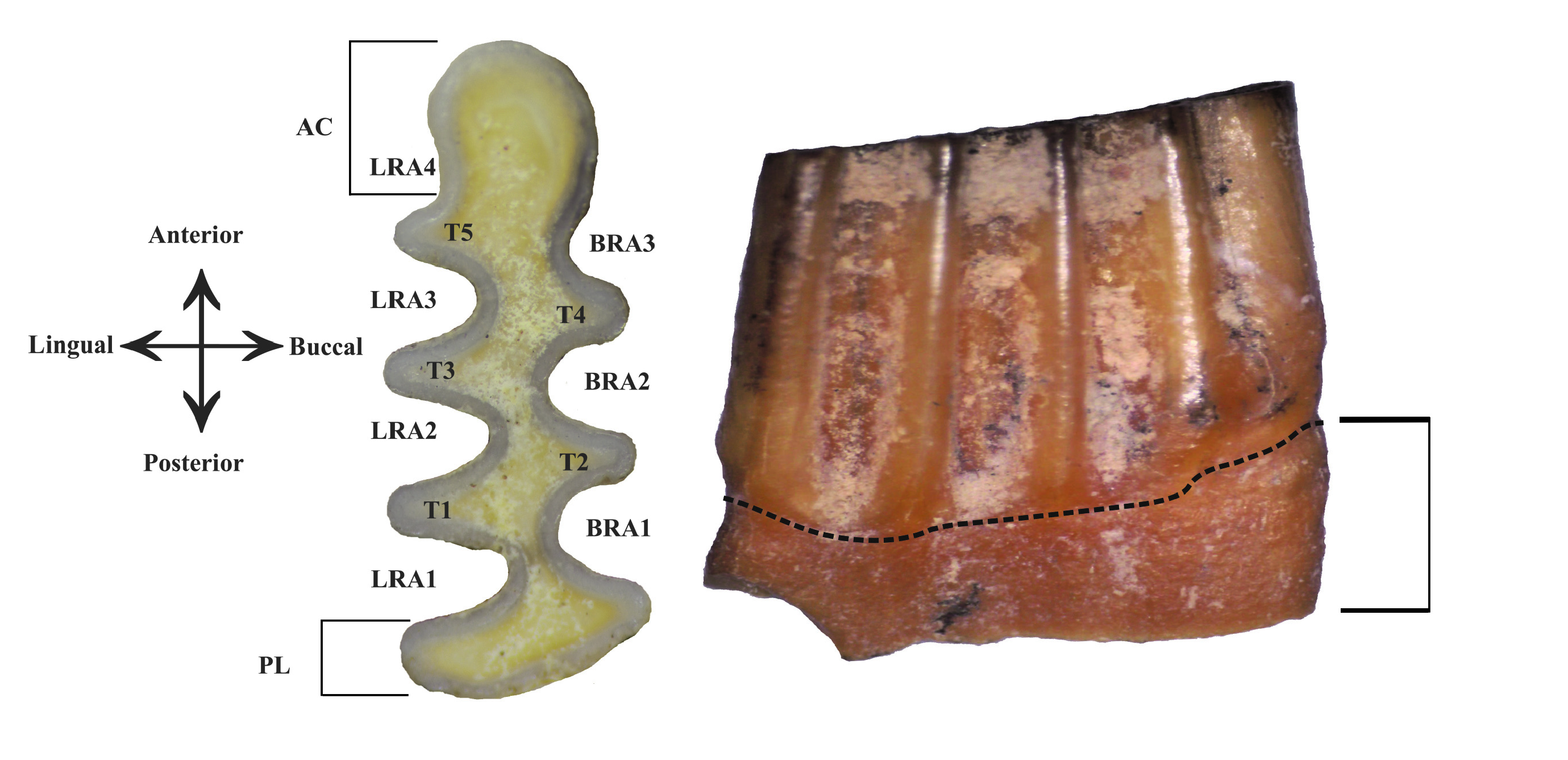
We restricted our analysis to the first lower molar, the most diagnostic tooth in arvicolines. The Ellobius lower m1 is composed of the anterior cap ( AC), five triangles ( T) with three buccal ( BRA) and four lingual ( LRA) re-entrant angles, and one posterior lobe ( PL) ( Figure 1A View FIGURE 1 ). Ellobius molars are notably characterized by broadly confluent triangles, and the presence of roots that are visible in adult and old individuals ( Figure 1B View FIGURE 1 ). Moreover, Ellobius molars lack cement in the re-entrant angles ( Coşkun, 2016).
For modern representatives, the skull morphology ( Kaya et al., 2018) and external characters (Kryštufek and Vohralík, 2009) contain the main diagnostic features, whereas fossil samples mostly consist of isolated molars or broken jaws. The occlusal morphology of the lower m1 is rather similar in the various Ellobius species (especially the three Iranian species E. fuscocapillus , E. lutescens and E. talpinus ). However, some specific morphological characters have been pointed out in previous studies: the AC is broad in Ellobius lutescens , narrow in Ellobius talpinus and elongated in Ellobius fuscocapillus ( Maul et al., 2015) ; the distance between T 4 and T 5 (W) and the total length (L) differ between the species, Ellobius fuscocapillus showing the largest teeth and Ellobius talpinus the smallest ( Rey-Rodríguez et al., 2020). However, these varying morphological and biometric characters are not always clear nor reliable distinction is possible.
| AC |
Amherst College, Beneski Museum of Natural History |
| T |
Tavera, Department of Geology and Geophysics |
| BRA |
Slovak National Museum |
| PL |
Západoceské muzeum v Plzni |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.